При переходе от состояния покоя к интенсивной мышечной деятельности во много раз возрастает потребность в кислороде. Скорость доставки кислорода является одним из важнейших факторов, определяющих возможности энергообеспечения работающих мышц.
Кислород воздуха через стенки легочных альвеол и кровеносных капилляров попадает в кровь путем диффузии вследствие разницы парциального давления в альвеолярном воздухе и крови. Парциальное давление кислорода в альвеолярном воздухе составляет 100 - 106 мм рт. ст., а в крови, притекающей к легким в покое, - 70 - 80 мм рт. ст., при мышечной работе парциальное давление кислорода в крови еще ниже.
186
Большая часть вдыхаемого кислорода связывается в эритроцитах с гемоглобином, который превращается в оксигемоглобин; причем, как было показано выше, каждая молекула гемоглобина способна связать четыре молекулы кислорода:
| Нb + 4О2 |
→ |
Нb4О2 |
| Гемоглобин |
|
Окскигемоглобин |
В крови взрослого человека содержится около 16 г гемоглобина. Подсчитано, что 100 г гемоглобина могут связать 134 мл кислорода (при 0°С и при атмосферном давлении), отсюда нетрудно определить кислородную емкость крови - общее количество связанного кровью кислорода. Она составляет 21 - 22 мл кислорода на 100 мл крови (при условии полного насыщения крови кислородом). На способность гемоглобина связывать кислород оказывает влияние температура и рН крови: чем ниже температура и выше рН, тем больше кислорода может связать гемоглобин.
Обогащенная кислородом кровь поступает в большой круг кровообращения. Сердце в покое перекачивает 5 - 6 л крови в минуту, следовательно, переносит от легких к тканям 250 - 300 мл кислорода. Во время интенсивной мышечной работы объем переносимой крови возрастает до 30 - 40 л/мин, а количество переносимого кровью кислорода - до 5 - 6 л/мин, т.е. увеличивается в 20 раз.
Увеличение содержания углекислого газа и повышение температуры крови в капиллярах мышечного волокна создают условия для освобождения кислорода из оксигемоглобина. Поскольку концентрация свободного кислорода в тканевых капиллярах выше, чем во внутриклеточном пространстве, происходит его диффузия в мышечные клетки, где обмен кислорода осуществляет миоглобин. Миоглобин связывает кислород и переносит его к митохондриям, где он используется в процессах, протекающих в аэробных условиях. Кроме того, миоглобин может депонировать кислород, а при интенсивной мышечной работе - отдавать свой кислородный запас.
При равномерной работе, если частота сердечных сокращений (ЧСС) не превышает 150 ударов в минуту, скорость потребления кислорода возрастает до тех пор, пока не наступит устойчивое состояние метаболических процессов, при котором потребление кислорода достигает постоянного уровня и в каждый данный момент времени точно соответствует потребности организма в нем. Такое устойчивое состояние называется истинным. Химически оно
187
характеризуется резким преобладанием дыхательного ресинтеза АТФ над анаэробным. При более интенсивной работе (ЧСС - 150- 180 ударов в минуту) не наблюдается установления устойчивого состояния, и потребление кислорода может возрастать до достижения максимального потребления (МПК). При достижении МПК может наблюдаться ложное устойчивое состояние, характеризующееся тем, что некоторое время потребление кислорода поддерживается на постоянном (максимальном) уровне. Это происходит не потому, что потребность организма в кислороде полностью удовлетворяется, а потому, что исчерпаны возможности сердечнососудистой системы по доставке кислорода к тканям. Максимальный уровень потребления кислорода не может поддерживаться долгое время. При длительной работе он снижается из-за утомления. Остановимся на определениях некоторых терминов, которые будем использовать при дальнейшем изложении материала.
- Кислородный запрос - количество кислорода, которое необходимо организму для полного удовлетворения энергетических потребностей за счет аэробных процессов.
- Кислородный приход - реальное потребление кислорода при интенсивной мышечной деятельности.
- Кислородный дефицит - разность между кислородным запросом и кислородным приходом.
Как видно из определений, кислородный приход всегда меньше кислородного запроса; в этом и состоит причина кислородного дефицита организма. В условиях кислородного дефицита происходит активация анаэробных процессов ресинтеза АТФ, что приводит к накоплению в организме продуктов анаэробного обмена. При установлении устойчивого состояния уровень метаболитов анаэробного обмена может снизиться за счет аэробных реакций; оставшаяся часть метаболитов устраняется в восстановительный период. Подводя итог сказанному, можно констатировать, что степень обеспечения организма кислородом - важнейший регулятор путей ресинтеза АТФ, расходуемой при мышечной деятельности.
В физиологии спорта принято различать и подразделять мышечную деятельность по зонам мощности: максимальная, субмаксимальная, высокая и умеренная. Существует и другое подразделение работы: в анаэробной, в смешанной и в аэробной зоне энергообеспечения.
Во всякой мышечной работе прежде всего следует различать начальную (пусковую) ее фазу и следующее за тем продолжение. Время пусковой фазы зависит от интенсивности работы: чем интенсивнее работа, тем продолжительнее пусковая фаза и тем резче
188
выражены вызываемые ею биохимические изменения в мышцах. В первые секунды работы мышцы получают меньше кислорода, чем им необходимо. Создавшийся кислородный дефицит тем больше, чем выше интенсивность работы, чем в большей мере возрастает потребность в кислороде (кислородный запрос). Поэтому в пусковой фазе ресинтез АТФ происходит исключительно анаэробными путями (креатинкиназная реакция, гликолиз).
Если интенсивность мышечной работы максимальна (а длительность, естественно, кратковременна), то на этой пусковой фазе она и заканчивается; следовательно, кислородный запрос будет неудовлетворен.
При работе субмаксимальной интенсивности, но большей длительности, биохимические изменения в пусковой фазе станут менее резки, а сама пусковая фаза укоротится. Потребление кислорода достигает предельно возможных величин (МПК), но и этих количеств кислорода недостаточно для удовлетворения кислородного запроса организма, который очень велик; в этих условиях организм испытывает кислородный дефицит. Значение креатинкиназного пути уменьшится, гликолиз будет еще достаточно интенсивен, но в известной мере начнет включаться и дыхательное регенерирование АТФ. Субстратом гликолиза будет не столько глюкоза, полученная от распада гликогена мышц, сколько глюкоза, приносимая кровью из печени.
При мышечной деятельности еще меньшей интенсивности и большей длительности после кратковременной пусковой фазы преобладающее значение получает ресинтез АТФ по аэробному механизму, поскольку постепенно устанавливается равновесие между кислородным запросом и кислородным приходом. Уровень АТФ в мышцах повышается (но не до исходных величин) и стабилизируется; повышается и уровень креатинфосфата, но в меньшей степени, чем АТФ.
Если при продолжении мышечной работы резко увеличить ее мощность, то в известной мере повторится то, что наблюдалось в пусковой фазе. Поскольку увеличение мощности работы влечет за собой и увеличение кислородного запроса, а он не может быть моментально удовлетворен, в энергообеспечение мышечной деятельности снова включатся анаэробные механизмы ресинтеза АТФ.
И еще раз рассмотрим последовательность включения различных путей ресинтеза АТФ уже с позиций удовлетворения потребности организма в кислороде: первые 2 - 3 сек энергообеспечение мышечной деятельности осуществляется за счет расщепления АТФ мышц; затем начинается ее ресинтез (от 3 до 20 сек) -
189
преимущественно за счет расщепления креатинфосфата, через 30 - 40 сек максимальной интенсивности достигает гликолиз; далее постепенно все больше превалирует аэробный механизм ресинтеза АТФ - окислительное фосфорилирование (рис. 38).
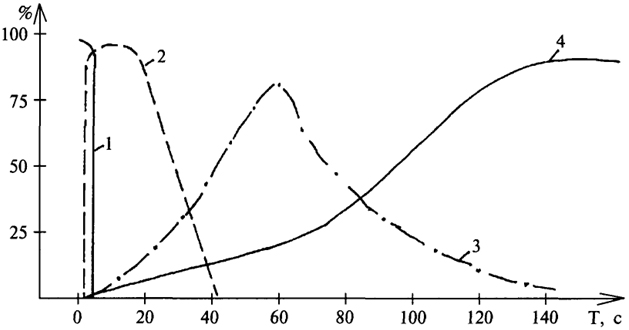
Рис. 38. Участие различных источников энергии в энергообеспечении мышечной деятельности в зависимости от ее длительности (
по Н.Н. Яковлеву, 1983): 1 - расщепление АТФ; 2 - распад Кф; 3 - гликолиз; 4 - аэробное окисление
Мощность аэробного энергообразования оценивается величиной МПК. В таблице 14 представлены МПК спортсменов и спортсменок различных специальностей, при анализе которых можно сделать заключение о вкладе аэробного механизма ресинтеза АТФ в энергообеспечение мышечной деятельности в процессе выполнения различных по продолжительности и мощности упражнений.
Таблица 14
МПК спортсменов и спортсменок (мл/мин·кг)
| Вид спорта |
Спортсмены |
Спортсменки |
| Лыжи |
80 |
60 |
| Бег (3000 м) |
78 |
- |
| Бег (800 - 1500 м) |
70 |
62 |
| Велосипед |
69 - 70 |
64 |
| Горные лыжи |
63 |
54 |
| Плавание |
61 |
52 |
| Гимнастика |
56 |
43 |
| Нетренированные люди |
42 - 43 |
37 - 38 |
190
Систематическая физическая нагрузка приводит к увеличению числа и относительного объема митохондрий в мышечной клетке, а также к существенным изменениям в их внутренней мембране: в ней увеличивается количество крист и составляющих их ансамблей дыхательных ферментов; повышается активность дыхательных ферментов, что создает преимущества тренированному организму в отношении более полного использования поступающего в клетки кислорода и накопления энергии.
191
