В тканях организма происходит непрерывное обновление липидов. Основную массу липидов тела человека составляют триглицериды, которыми особенно богата жировая ткань. В виде включений триглицериды имеются в большинстве тканей и органов. Поскольку липиды выполняют энергетическую функцию, то процессы их обновления связаны с мобилизацией и депонированием их в процессе образования энергии. Обмен фосфолипидов недостаточно изучен, но имеется мнение, что их обновление связано прежде всего с процессами восстановления структуры мембран.
114
Обновление липидов тканей и органов организма требует предварительного внутриклеточного ферментативного их гидролиза.
Гидролиз триглицеридов проходит в два этапа. На первом этапе происходит гидролиз внешних сложноэфирных связей, ускоряет этот процесс фермент липаза. В клетках организма человека функционирует несколько видов липаз, имеющих разную локализацию и оптимум рН. В лизосомах локализованы кислые липазы (липазы, проявляющие максимальную активность в кислой среде), в цитоплазме - нейтральные, в микросомах - щелочные. Активация липаз происходит по механизму фосфорилирования - дефосфорилирования, как и у гликогенфосфорилазы. Гидролиз триглицеридов называется липолизом.
Первый этап липолиза можно представить в виде схемы реакции:

β-Моноглицерид, образовавшийся на первом этапе распада триглицеридов, далее гидролизуется неспецифической эстеразой до глицерина и высшей жирной кислоты:
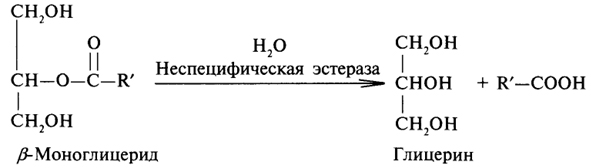
В результате гидролиза триглицеридов образуется глицерин и три молекулы высших жирных кислот.
Гидролиз фосфатидов. Фосфатиды распадаются на соответствующие структурные компоненты: глицерин, ВЖК, фосфорную кислоту и азотистое основание. Процессы гидролиза сложноэфирных связей в молекуле фосфатидов ускоряются различными по
115
специфичности фосфолипазами. В зависимости от того, гидролиз какой сложноэфирной связи катализирует фосфолипаза, ее называют А1, A2, С, D (схема 5).
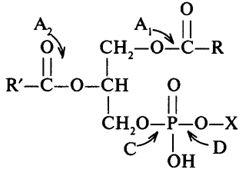
Схема 5. Специфичность действия фосфолипаз: X - азотистое основание; стрелки указывают гидролизуемую соответствующей фосфолипазой связь
Продукты гидролиза триглицеридов и фосфатидов подвергаются дальнейшим метаболическим превращениям.
Обмен глицерина может осуществляться несколькими путями. Значительная часть образовавшегося при гидролизе липидов глицерина используется для ресинтеза триглицеридов. Второй путь обмена глицерина - включение продукта его окисления в гликолиз или в глюконеогенез. Независимо от пути обмена начальным этапом является процесс фосфорилирования глицерина, донором фосфатной группы является молекула АТФ:

Большая часть α-глицерофосфата используется для синтеза триглицеридов. Обмен глицерина тесно связан с гликолизом, во второй этап которого вовлекаются его метаболиты:

116
Окисление жирных кислот. В 1904 г. Ф. Кнооп показал, что в митохондрии в ходе окисления происходит постепенное уменьшение на два углеродных атома с карбоксильного конца высшей жирной кислоты. Ф. Кнооп назвал механизм окисления ВЖК β-окислением. Дальнейшие исследования, проведенные А. Ленинджером, Ф. Линеном, Д. Грином, С. Очоа и другими учеными, уточнили и развили представления о β-окислении высших жирных кислот.
Первым этапом распада жирных кислот является их активирование; этот процесс катализируется ацил-КоА-синтетазой, которая локализована в мембранах эндоплазматической сети и в наружной мембране митохондрий:
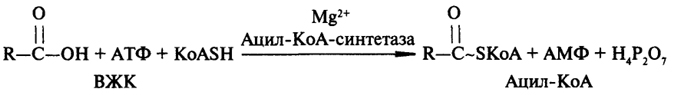
Поскольку процесс активирования ВЖК идет вне митохондрий, то далее необходим транспорт ацила через мембрану внутрь митохондрий. Транспорт происходит с участием находящегося на внешней стороне мембраны карнитина, на который передается ацил с ацил-КоА из цитоплазмы клетки. Затем ацилкарнитин диффундирует через мембрану митохондрии и передает свой ацил коэнзиму А, находящемуся в матриксе митохондрии. Перенос ацила между КоА и карнитином является ферментативным процессом, катализируемым ацил-КоА-карнитин-трансферазой (рис. 16).
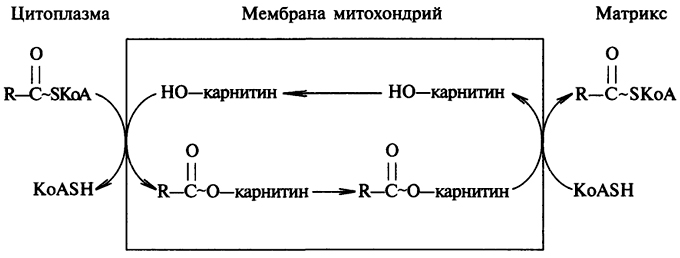
Рис. 16. Схема транспорта жирных кислот через митохондриальную мембрану
В матриксе митохондрии происходит процесс β-окисления. Первой стадией β-окисления ВЖК является окисление ацил-КоА путем отщепления двух атомов водорода от α- и β-углеродных атомов ацила коферментом соответствующей дегидрогеназы:
117

Далее происходит присоединение молекулы воды таким образом, что ОН-группа присоединяется к β-, а атом водорода - к α-углеродному атому:
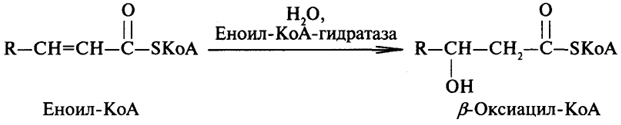
На следующей стадии идет окисление β-оксиацил-КоА, катализируемое дегидрогеназой, в результате действия которой образуется β-кетоацил-КоА:

На последней стадии β-окисления происходит негидролитический распад β-кетоацил-КоА и перенос ацила, укороченного на два углеродных атома по сравнению с первоначальным, на молекулу КоА:

Ацил-КоА, образовавшийся на последнем этапе, вновь подвергается β-окислению, проходя все описанные выше стадии. Следовательно β-окисление - это циклический процесс. Конечным продуктом β-окисления высших жирных кислот является ацетил-КоА, дальнейший обмен которого зависит от состояния организма. Однако в какой бы путь обмена он не вступал, результатом будет освобождение КоА, запасы которого в клетке ограничены. β-Окисление ВЖК является одним из основных источников
118
получения энергии для синтеза АТФ в животной клетке. В главе "Основы биоэнергетики" будет подробно рассмотрен энергетический эффект этого процесса.
Образование кетоновых тел. Одним из процессов, в котором происходит регенерирование свободного КоА из его ацильных производных, является образование ацетоуксусной кислоты. В этом процессе принимают участие три молекулы ацетил- КоА. Сначала происходит конденсация двух молекул ацетил-КоА с образованием β-кетобутирил-КоА:
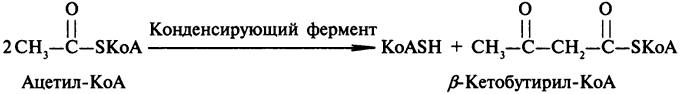
На второй стадии происходит высвобождение КоА из β-кетобутирил-КоА. Для этого процесса нужна еще одна молекула ацетил-КоА:
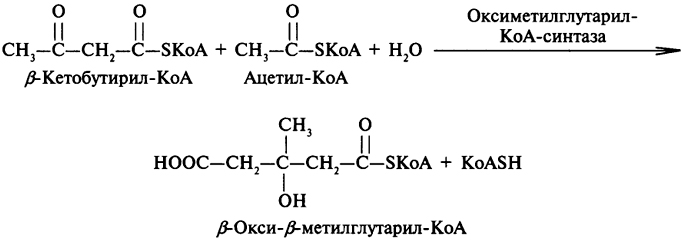
Образовавшийся β-окси-β-метилглутарил-КоА далее подвергается негидролитическому расщеплению, в результате образуется ацетоуксусная кислота:

119
В результате конденсации трех молекул ацетил-КоА образуется молекула ацетоуксусной кислоты и высвобождаются две молекулы КоА.
Ацетоуксусная кислота при восстановлении дает β-оксимасляную кислоту:

Ацетоуксусная и β-оксимасляная кислоты синтезируются в печени и поступают с кровью к мышечной и другим тканям, которые утилизируют их в цикле Кребса. Нарушения в обмене жиров сопровождаются накоплением ацетоуксусной и β-оксимасляной кислот в крови. Ацетоуксусная кислота может превращаться в ацетон по схеме:
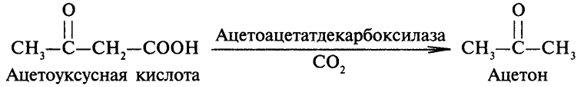
Ацетоуксусная, β-оксимасляная кислоты и ацетон получили название кетоновых тел. Усиленное образование их называется кетозом. Состояние организма, при котором происходит избыточное накопление кетоновых тел в крови, называют кетонемией, а выделение их с мочой - кетонурией. Среди многих причин патологического накопления кетоновых тел особенно важными считают дефицит поступающих с пищей углеводов (относительно окисляющихся липидов) и нарушение обмена углеводов и жирных кислот при недостатке инсулина.
Основными структурными блоками триглицеридов и фосфатидов являются α-глицерофосфат и ацильные производные КоА (ацил-КоА). α-Глицерофосфат образуется из глицерина, возникающего при распаде глицеринсодержащих липидов, а высшие жирные кислоты синтезируются из малонил-КоА. Рассмотрим подробно процесс биосинтеза ВЖК.
Синтез высших жирных кислот локализован в эндоплазматической сети клетки. Непосредственным источником синтеза является
120
малонил-КоА, образующийся из ацетил-КоА и оксида углерода (IV) при участии АТФ:
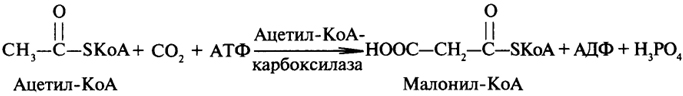
Следует подробнее остановиться на характеристике ацетил-КоА-карбоксилазы, поскольку она является полифункциональным ферментом. Ацетил-Ко А-карбоксилаза представляет собой полипептидную цепь, имеющую доменную структуру; каждый домен в полифункциональном ферменте обладает определенной каталитической активностью. Некоторые домены в своем составе могут иметь кофакторы (коферменты). Ацетил-КоА-карбоксилаза содержит домен биотинкарбоксилазы, биотин-карбоксилпроводящий домен и домен транскарбоксилазы. Все три домена согласованно ускоряют синтез малонил-КоА, который поступает на второй полифункциональный фермент - синтетазу высших жирных кислот, - при посредстве которого и происходит синтез ВЖК.
В составе синтетазы ВЖК выделяют три домена, каждый из которых несет определенную функциональную нагрузку. Первый домен отвечает за элонгацию цепи, второй - за восстановление цепи ВЖК, третий - за высвобождение синтезированного ацила ВЖК из комплекса с ферментом в виде ацила-КоА. Рассмотрим процесс синтеза ВЖК, выделяя работу каждого домена.
Начальным этапом синтеза ВЖК является конденсация малонил-КоА с ацетил-КоА:

Образовавшийся β-кетобутирил-КоА сначала восстанавливается до β-оксибутирил-КоА, который далее с участием дегидратазы превращается в кротонил-КоА, содержащий двойную связь. Кротонил-КоА восстанавливается до бутирил-КоА.
Следует подчеркнуть, что ферменты редуктазы в своем составе содержат НАДФ. Работу второго домена синтетазы ВЖК можно представить следующей цепью превращений:
121
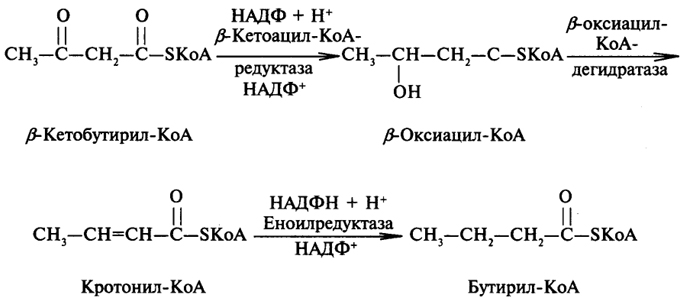
Биосинтез ВЖК носит циклический характер. Синтезированный бутирил-КоА вступает в новый цикл превращений, представленных выше. Для синтеза, например, пальмитиновой кислоты нужно семь таких циклов; в каждом цикле происходит удлинение ацила на два углеродных атома. По достижении ацильным радикалом длины в 16 и более атомов углерода происходит его отщепление от фермента третьим доменом, обладающим тиоэстеразной активностью.
Синтез триглицеридов происходит при депонировании липидов в жировой или других тканях организма. Локализован этот процесс на мембранах эндоплазматической сети.
Первой стадией синтеза триглицеридов является трансацилирование α-глицерофосфата с образованием фосфатидной кислоты:

Далее фосфатидная кислота подвергается действию фосфатидат-фосфатазы с образованием α, β-диацилглицерина:
122

На последней стадии синтеза происходит ацилирование свободной ОН-группы α; β-диацилглицерина, катализируемое диацилглицерин-ацилтрансферазой:

Ферменты, ускоряющие синтез триглицеридов, найдены в клетках печени, слизистой оболочки кишечника, жировой ткани и др. Из тканей с интенсивным синтезом триглицеридов, они мигрируют в ткани, где нет активного синтеза.
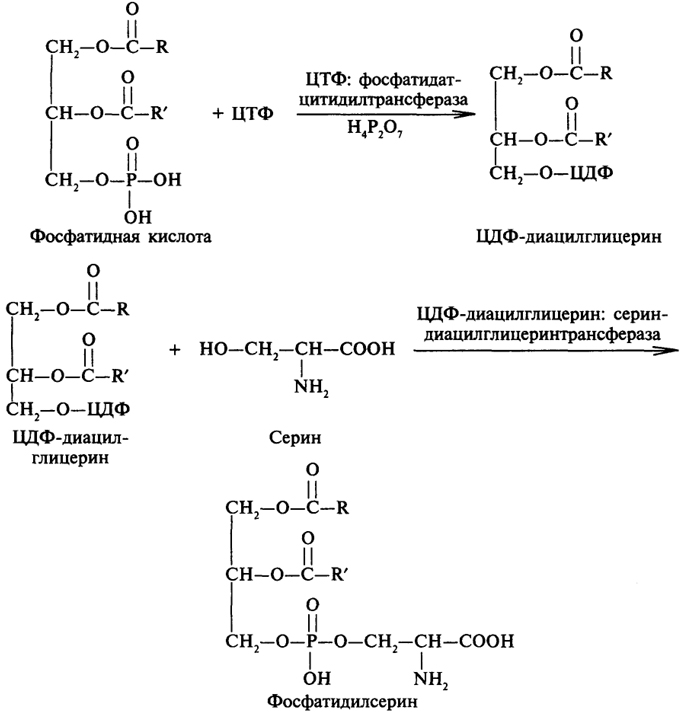
Синтез фосфатидов. Существует два пути синтеза фосфатидов, причем для обоих необходима цитидинтрифосфорная кислота (ЦТФ). Ниже представлен путь синтеза фосфатидов из фосфатидной кислоты, синтез которой был рассмотрен выше.
Взаимодействие фосфатидной кислоты с ЦТФ приводит к образованию ЦДФ-диацилглицерина, который как кофермент способен участвовать в переносе диацилглицерина на азотистое основание, например серии. В результате этого превращения образуется фосфатидилсерин, который можно рассматривать в качестве исходного соединения для образования другого фосфатида - фосфатидилэтаноламина:
123
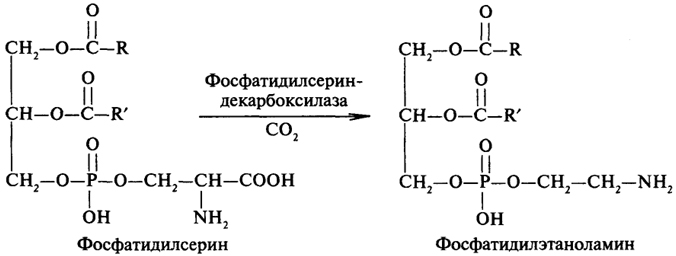
Декарбоксилирование фосфатидилсерина приводит к образованию фосфатидилэтаноламина:
124
