Первичная и вторичная продукции. Скорость, с которой продуценты экосистемы фиксируют солнечную энергию в химических связях синтезируемого органического вещества, определяет продуктивность сообществ. Органическую массу, создаваемую растениями за единицу времени, называют первичной продукцией сообщества. Продукцию выражают количественно в сырой или сухой массе растений либо в энергетических единицах - эквивалентном числе джоулей.
Валовая первичная продукция - количество вещества, создаваемого растениями за единицу времени при данной скорости фотосинтеза. Часть этой продукции идет на поддержание жизнедеятельности самих растений (траты на дыхание). Эта часть может быть довольно большой. В тропических лесах и зрелых лесах умеренного пояса она составляет от 40 до 70% валовой продукции. Планктонные водоросли используют на метаболизм около 40% фиксируемой энергии. Такого же порядка траты на дыхание у большинства сельскохозяйственных культур. Оставшаяся часть созданной органической массы характеризует чистую первичную продукцию, которая представляет собой величину прироста растений. Чистая первичная продукция - это энергетический резерв для консументов и редуцентов. Перерабатываясь в цепях питания, она идет на пополнение массы гетеротрофных организмов.
Прирост за единицу времени массы консументов - это вторичная продукция сообщества. Вторичную продукцию вычисляют отдельно для каждого трофического уровня, так как прирост массы на каждом из них происходит за счет энергии, поступающей с предыдущего.
Гетеротрофы, включаясь в трофические цепи, живут в конечном итоге за счет чистой первичной продукции сообщества.
48
В разных экосистемах они расходуют ее с разной полнотой. Если скорость изъятия первичной продукции в цепях питания отстает от темпов прироста растений, то это ведет к постепенному увеличению общей биомассы продуцентов. Под биомассой понимают суммарную массу организмов данной группы или всего сообщества в целом. Часто биомассу выражают в эквивалентных энергетических единицах.
Недостаточная утилизация продуктов опада в цепях разложения имеет следствием накопление в системе мертвого органического вещества, что происходит, например, при заторфовывании болот, зарастании мелководных водоемов, создании больших запасов подстилки в таежных лесах и т. п. Биомасса сообщества с уравновешенным круговоротом веществ остается относительно постоянной, так как практически вся первичная продукция тратится в цепях питания и разложения.
Правила пирамид. Экосистемы очень разнообразны по относительной скорости создания и расходования как чистой первичной продукции, так и чистой вторичной продукции на каждом трофическом уровне. Однако всем без исключения экосистемам свойственны определенные количественные соотношения первичной и вторичной продукции, получившие название правило пирамиды продукции: на каждом предыдущем трофическом уровне количество биомассы, создаваемой за единицу времени, больше, чем на последующем. Графически это правило часто выражают в виде пирамид, суживающихся кверху и образованных поставленными друг на друга прямоугольниками равной высоты, длина которых определяет масштабы продукции на соответствующих трофических уровнях. Пирамида продукции отражает законы расходования энергии в пищевых цепях.
Скорость создания органического вещества не определяет его суммарные запасы, т.е. общую биомассу всех организмов каждого трофического уровня. Наличная биомасса продуцентов или кон-сументов в конкретных экосистемах зависит от того, как соотносятся между собой темпы накопления органического вещества на определенном трофическом уровне и передачи его на вышестоящий, т.е. насколько сильно выедание образовавшихся запасов. Немаловажную роль при этом играет скорость оборота генераций основных продуцентов и консументов.
В большинстве наземных экосистем действует также правило пирамиды биомасс: суммарная масса растений оказывается больше, чем биомасса всех фитофагов и травоядных, а масса тех, в свою очередь, превышает массу всех хищников. Отношение годового прироста растительности к биомассе в наземных экосистемах сравнительно невелико. В разных фитоценозах, где основные продуценты различаются по длительности жизненного цикла, размерам и темпам роста, это соотношение варьирует от 2 до 76%. Особенно
49
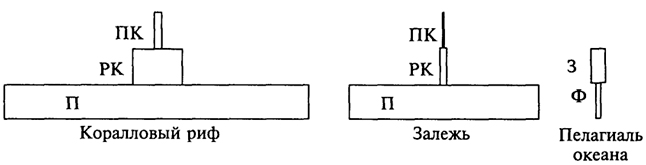
Рис. 3.2. Пирамиды биомассы некоторых биоценозов: П - продуценты; РК - растительноядные консументы; ПК - плотоядные консументы; Ф - фитопланктон; 3 - зоопланктон (крайняя справа пирамида биомассы имеет перевернутый вид)
низки темпы относительного прироста биомассы в лесах разных зон, где годовая продукция составляет лишь 2 - 6 % от общей массы растений, накопленной в телах долгоживущих крупных деревьев. Даже в наиболее продуктивных дождевых тропических лесах эта величина не превышает 6,5%. В сообществах с господством травянистых форм скорость воспроизводства биомассы гораздо выше: годовая продукция в степях составляет 41-55%, а в травяных тугаях и эфемерно-кустарниковых полупустынях достигает даже 70 - 76%. Пирамиды биомасс некоторых биоценозов приведены на рис. 3.2. Иногда эти пирамиды изображаются иначе (рис. 3.3).
Отношение первичной продукции к биомассе растений определяет масштабы выедания растительной массы, которые возможны в сообществе без подрыва его продуктивности. Относительная доля потребляемой животными первичной продукции в травянистых сообществах выше, чем в лесах. Копытные, грызуны, насекомые-фитофаги в степях используют до 70% годового прироста растений, тогда как в лесах в среднем не более 10%. Однако возможные пределы отчуждения растительной массы животными в наземных сообществах не реализуются полностью и значительная часть ежегодной продукции поступает в опад.
В океанах, где основными продуцентами являются одноклеточные водоросли с высокой скоростью оборота генераций, их годовая продукция в десятки и даже сотни раз может превышать запас биомассы. Вся чистая первичная продукция так быстро вовлекается в цепи питания, что накопление биомассы водорослей очень мало, но вследствие высоких темпов размножения небольшой их запас оказывается достаточным для поддержания скорости воссоздания органического вещества (рис. 3.4).
Для океана правило пирамиды биомасс недействительно, она имеет перевернутый вид (см. рис. 3.2). На высших трофических уровнях преобладает тенденция к накоплению биомассы, так как длительность жизни крупных хищников велика, скорость оборота их генераций, наоборот, мала, и в их телах задерживается значительная часть вещества, поступающего по цепям питания.
50

Рис. 3.3. Пирамида биомасс и трофические уровни в экосистеме. Биомасса дана в условных числах
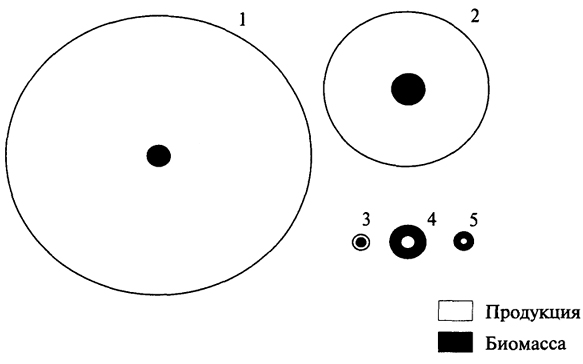
Рис. 3.4. Схема соотношения продукции и биомассы у бактерий (1), фитопланктона (2), зоопланктона (3), бентоса (4) и рыб (5) в Баренцевом море
51
В тех трофических цепях, где передача энергии происходит в основном через связь "хищник-жертва", часто выдерживается правило пирамиды чисел: общее число особей, участвующих в цепях питания, с каждым звеном уменьшается. Это связано с тем, что хищники, как правило, крупнее объектов своего питания и для поддержания биомассы одного хищника нужно несколько или много жертв. Из этого правила могут быть и исключения - те редкие случаи, когда более мелкие хищники живут за счет групповой охоты на крупных животных. Правило пирамиды чисел было подмечено еще в 1927 г. Ч. Элтоном, который отметил также, что оно неприменимо к цепям питания паразитов, размеры которых с каждым звеном уменьшаются, а число особей возрастает.
Все три правила пирамид - продукции, биомассы и чисел - выражают в конечном итоге энергетические отношения в экосистемах, и если первые два проявляются в сообществах с определенной трофической структурой, то последнее (пирамида продукции) имеет универсальный характер.
Знание законов продуктивности экосистем, возможность количественного учета потока энергии имеют чрезвычайное практическое значение. Первичная продукция агроценозов и эксплуатации человеком природных сообществ - основной источник запасов пищи для человечества. Не менее важна и вторичная продукция, получаемая за счет сельскохозяйственных и промышленных животных, так как животные белки включают целый ряд незаменимых для людей аминокислот, которых нет в растительной пище. Точные расчеты потока энергии и масштабов продуктивности экосистем позволяют регулировать в них круговорот веществ таким образом, чтобы добиваться наибольшего выхода выгодной для человека продукции. Кроме того, необходимо хорошо представлять допустимые пределы изъятия растительной и животной биомассы из природных систем, чтобы не подорвать их продуктивность. Подобные расчеты обычно очень сложны из-за методических трудностей и точнее всего выполнимы для более простых водных экосистем.
Распределение биологической продукции. Важнейшим практическим результатом энергетического подхода к изучению экосистем явилось осуществление исследований по Международной биологической программе, проводившихся учеными разных стран мира начиная с 1969 г. в целях изучения потенциальной биологической продуктивности Земли.
Теоретическая возможная скорость создания первичной биологической продукции определяется возможностями фотосинтетического аппарата растений. Максимально достигаемый в природе КПД фотосинтеза 10 -12 % энергии ФАР, что составляет около половины от теоретически возможного. Такая скорость связывания энергии достигается, например, в зарослях джугары и тростника
52
в Таджикистане в кратковременные, наиболее благоприятные периоды. КПД фотосинтеза 5 % считается очень высоким для фитоценоза. В целом по земному шару усвоение растениями солнечной энергии не превышает 0,1 %, так как фотосинтетическая активность растений ограничивается множеством факторов.
Обшая годовая продукция сухого органического вещества на Земле составляет 150 - 200 млрд т. Около 1/3 его образуется в океанах, около 2/3 - на суше. Почти вся чистая первичная продукция Земли служит для поддержания жизни всех гетеротрофных организмов. Энергия, недоиспользованная консументами, запасается в их телах, органических осадках водоемов и гумусе почв (табл. 1,2).
Благоприятное сочетание многих природных факторов приводит к тому, что наиболее продуктивной наземной экосистемой являются тропические леса. Во влажных экваториальных лесах встречается более 50000 видов растительности. Биомасса экваториального леса достигает 1000 т/га (в бассейне Амазонки 1700 т/га) при годовом приросте 50 т/га. Господствующим компонентом этих лесов являются деревья, составляющие около 70 % всех встречающихся видов внешних растений. В лесах Амазонки можно встретить на площади 1 га от 50 до 100 видов растений. Таким же богатством и разнообразием отличается животный мир влажных экваториальных лесов.
Эффективность связывания растительностью солнечной радиации снижается при недостатке тепла и влаги, при неблагоприятных физических и химических свойствах почвы и т.п. Продуктивность растительности изменяется не только при переходе от одной климатической зоны к другой, но и в пределах каждой зоны. На территории СССР в зонах достаточного увлажнения первичная продуктивность увеличивалась с севера на юг с ростом притока тепла и продолжительности вегетационного сезона. Годовой прирост растительности изменялся от 20 ц/га на побережье и островах Северного Ледовитого океана до более чем 200 ц/га на Черноморском побережье Кавказа.
Средний коэффициент использования энергии ФАР для всей территории СССР составлял 0,8%: от 1,8 - 2,0% на Кавказе до 0,1 -0,2 % в пустынях Средней Азии. В большинстве восточных районов страны, где условия увлажнения менее благоприятны, этот коэффициент составлял 0,4 - 0,8%, на европейской территории - 1,0-1,2%. КПД суммарной радиации примерно вдвое ниже.
Для пяти континентов мира средняя продуктивность различается сравнительно мало. Исключением является Южная Америка, на большей части которой условия для развития растительности очень благоприятны (табл. 3).
Питание людей обеспечивается в основном сельскохозяйственными культурами, занимающими примерно 10% площади суши
53
Таблица 1
Продуктивность основных экосистем земного шара
| Экосистемы |
Площадь,
млн км2 |
Продуктивность
растений, г/(м2.год) |
Общая
продукция
растений
млрд т/год |
Общая
продукция
животных
млрд т/год |
Крайние
значения |
Средние |
Материковые
экосистемы
(в целом) |
149 |
0 - 3500 |
773 |
115 |
909 |
| в том числе: |
Влажные
тропические леса |
17 |
1000 - 3500 |
2200 |
37,4 |
260 |
Вечнозеленые
леса умеренного
пояса |
5 |
600 - 2500 |
1300 |
6,5 |
26 |
Листопадные леса
умеренного пояса |
7 |
600 - 2500 |
1200 |
8,4 |
42 |
| Тайга |
12 |
400 - 2000 |
800 |
9,6 |
38 |
| Саванна |
15 |
200-2000 |
900 |
13,5 |
300 |
Тундры и
высокогорья |
8 |
10 - 400 |
140 |
1,1 |
3 |
Пустыни и
полупустыни |
18 |
10 - 250 |
90 |
1,6 |
7 |
Болота мангровые
и низинные |
2 |
800 - 3500 |
2000 |
4,0 |
32 |
| Озера и водостоки |
2 |
100 - 1500 |
250 |
0,5 |
10 |
Культивируемые
земли |
14 |
100 - 3500 |
650 |
9,1 |
9 |
Морские
экосистемы
(в целом) |
361 |
2 - 2400 |
152 |
55,0 |
3025 |
| в том числе: |
| Открытый океан |
332 |
2 - 400 |
125 |
41,5 |
2500 |
| Аппвелинги |
0,4 |
400 - 1000 |
500 |
0,2 |
11 |
Континентальный
шлейф |
26 |
200 - 600 |
360 |
9,6 |
430 |
Заросли
водорослей
и рифы |
0,6 |
500 - 4000 |
2500 |
1,6 |
36 |
| Эстуарии |
1,4 |
200 - 3500 |
1500 |
2,1 |
48 |
Средняя
продуктивность
биосферы |
510 |
0 - 4000 |
333 |
170,0 |
3934 |
54
Таблица 2
Продуктивность и биомасса экосистем материков и океанов земного шара
| Экосистемы |
Площадь, км2 |
Растения |
Продукция животных, млрд т/год |
Биомасса животных, млрд т |
| Продукция |
Биомасса |
Потребление животными, млрд т/год |
| т/год (га·год) |
Глобальная, млрд т/год |
т/га |
Глобальная, млрд т |
| Континентов |
149 |
7,73 |
115,0 |
123,0 |
1837,0 |
7,8 |
0,909 |
1,005 |
| Океана |
361 |
1,52 |
55,0 |
0,1 |
3,9 |
20,2 |
3,045 |
0,997 |
| Всего |
510 |
3,33 |
170,0 |
3,6 |
1840,9 |
28,0 |
3,934 |
2,002 |
| Возделываемые земли |
14 |
6,50 |
9,1 |
10,0 |
14 |
0,09 |
0,009 |
0,006 |
Таблица 3
Продуктивность естественного растительного покрова
| Части света |
Средняя продуктивность, ц/га |
КПД ФАР |
| Европа |
85 |
1,26 |
| Азия |
98 |
0,88 |
| Африка |
103 |
0,59 |
| Северная Америка |
82 |
0,94 |
| Южная Америка |
209 |
1,13 |
| Австралия и Океания |
86 |
0,44 |
(около 1,4 млрд га). Общий годовой прирост культурных растений составляет около 16% всей продуктивности суши, большая часть которой приходится на леса.
Примерно половина урожая идет непосредственно на питание людей, остальная часть - на корм домашним животным, используется в промышленности и теряется в отбросах. Современные данные о потреблении человеческим обществом первичной продукции Земли приведены в главе 13.
Растительная пища обходится людям энергетически дешевле, чем животная. Сельскохозяйственные площади при рациональном использовании и распределении продукции могли бы обеспечить растительной пищей примерно вдвое большее население Земли, чем существующее. Однако сельскохозяйственное производство нуждается в большой затрате труда и больших капиталовложениях. Особенно трудно обеспечить население вторичной продукцией. В рацион человека должно входить не менее 30 г белков в день. Имеющиеся
55
на Земле ресурсы, включая продукцию животноводства и результаты промысла на суше и в океане, могут обеспечить ежегодно лишь около 50 % потребностей современного населения Земли.
Существующие ограничения, накладываемые масштабами вторичной продуктивности, усиливаются несовершенством социальных систем распределения. Большая часть населения Земли находится, таким образом, в состоянии хронического белкового голодания, а значительная часть людей страдает также и от общего недоедания (см. главу 7).
Таким образом, увеличение биологической продуктивности экосистем и особенно вторичной продукции является одной из основных задач, стоящих перед человечеством.
Вырубка леса на локальных участках с оставлением части территории под коренными типами лесной растительности вызывает ускоренные сукцессии, исходные фитоценозы восстанавливаются за относительно короткий срок - немногие десятилетия. Сплошные же рубки на больших площадях, особенно если используется мощная корчевальная техника, полностью разрушают не только лесное сообщество, но и весь почвенный покров, эволюция которого шла тысячелетия. В этом случае сукцессии приводят к иным, упрощенным типам сообществ, и на месте лесов возникают пустоши, болота или другие малопродуктивные экосистемы.
Таким образом, сообщество не может одновременно сочетать два противоположных свойства: быть высокостабильным и давать большой запас чистой продукции, который можно было бы изымать без вреда для самого ценоза.
Сукцессии, протекающие при разложении органического вещества в почве, лежат в основе биологического круговорота. Это естественные регуляторные процессы, восстанавливающие нормальное состояние нарушенных почв. Такие проблемы века, как подрыв естественного плодородия в результате нарушения процессов образования гумуса, загрязнение среды ядохимикатами и органическими отходами, эрозия, "утомление почв" и другие отрицательные явления, возникли вследствие ослабления регуляторных возможностей почв.
Стабильность экосистем, надежность протекания биологического круговорота веществ основаны на видовом разнообразии и полноте сукцессии, возникших в результате преобразования сообществ на основе длительной эволюции видов.
56
