Продолговатый мозг, Medulla oblongata, уже у круглоротых представлен в достаточно дифференцированном виде. Являясь продолжением спинного мозга, он на значительном протяжении сохраняет свойственные последнему структурные особенности и форму. Изменение конфигурации и взаимного расположения структур продолговатого мозга у всех позвоночных происходит в дорсальных отделах в результате формирования IV желудочка мозга путем расщепления крыловидной пластинки и расхождения ее в вентролатеральных направлениях. Рострально желудочек переходит в полость среднего мозга. Верхняя его часть сформирована хориоидной крышей и у большинства позвоночных на всем протяжении ограничена мозжечком.
Основной объем ядер продолговатого мозга составляют центральные проекции V-XII пар черепномозговых нервов. В зависимости от уровня филогенетической или адаптивной эволюции выраженность этих отделов варьирует, что и обусловливает значительное расхождение во внешнем виде и топографии медуллярного отдела. При анализе строения продолговатого мозга традиционно рассматривают локализацию ядер по нескольким колоннам, параллельным оси мозга. Границы их образованы бороздами на стенках желудочка, дополняющими пограничную борозду в период развития мозга и сохраняющимися в ряде случаев у взрослых животных. Эти борозды разделяют серое вещество на зоны, которые являются продолжением соответствующих участков спинного мозга (рис. 26).
Соматическая афферентная зона представлена в продолговатом мозге соматосенсорной (система тройничного нерва и
64
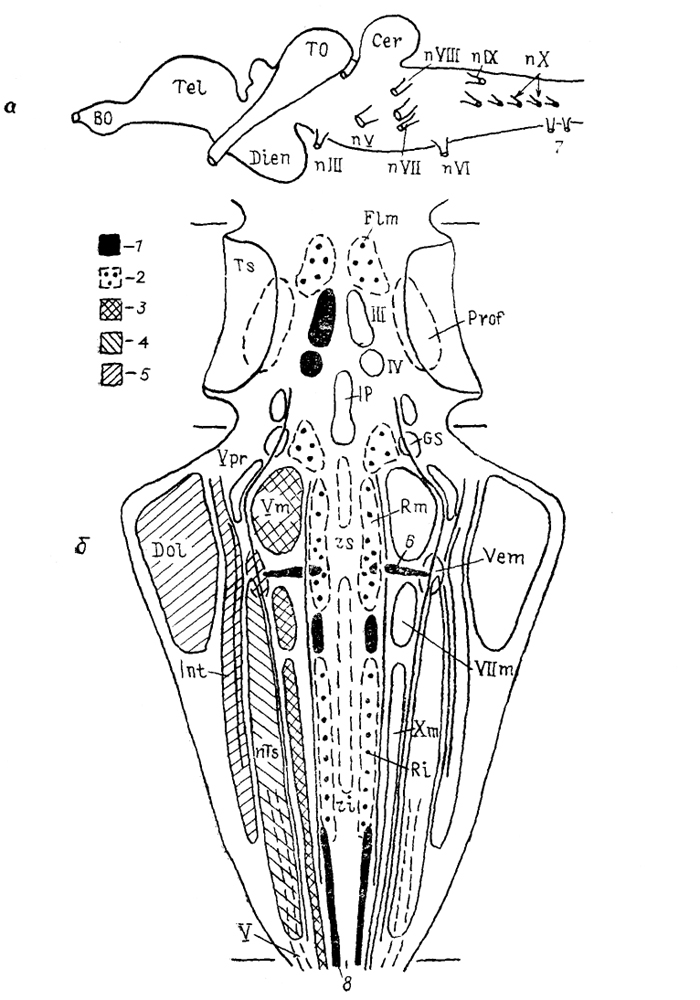
Рис. 26. Строение
(а) и расположение
(б) важнейших ядер стволовой части головного мозга анамний (Nieuwenhuys, Meek, 1985).
1, 3-5, 8 - колонны: соматомоторная (1), висцеромоторная (3), висцеросенсорная (4), специальная соматосенсорная (5), моторная спинного мозга (8); 2 - ретикулярная формация; 6 - маутнеровская клетка; 7 - спинно-окципитальные нервы.
65
ядра дорсальных канатиков) и специализированной октаво-латеральной (система вестибуло-кохлеарного нерва и в ряде случаев - водные позвоночные - часть лицевого нерва) областями. Висцеросенсорная зона содержит ядра языкоглоточного, лицевого и блуждающего нервов, связанные с обработкой информации от вкусовых и висцерохимических рецепторов.
Соматомоторная зона объединяет многочисленные клетки, составляющие на уровне продолговатого мозга ядра отводящего и подъязычного нервов. Эта колонна продолжается и на уровне среднего мозга, где она содержит ядра глазодвигательного и блокового нервов. Каудально она переходит в моторную колонну спинного мозга. Висцеромоторная зона образована ядрами тройничного, лицевого, языкоглоточного и блуждающего нервов.
Внутренняя топография того или иного отдела и его компонентов чрезвычайно вариабельна, и далеко не всегда структурная организация продолговатого мозга четко соответствует вышеизложенной. Так, анатомические границы зон часто не совпадают с границами колонн в силу различного развития структурных элементов, их образующих. Кроме того, во многих группах животных отмечается гипертрофия одних отделов и слабое развитие других. В результате специализация сенсорных систем, отражаясь в строении продолговатого мозга, приводит к значительным изменениям его конфигурации, причем у низших позвоночных эти вариации заметнее, чем у высших.
Вместе с тем наличие ярко выраженной специализации какой-либо системы оказывается не только интересной иллюстрацией адаптивных возможностей мозга, но и позволяет вычленить (при сравнительном анализе) базовые признаки, характеризующие те или иные отделы мозга. В этом отношении одной из наиболее интересных групп является надкласс Рыбы, где в зависимости от условий сред обитания и соответствующей специализации сенсорных и моторных систем обнаруживается чрезвычайно широкий диапазон вариантов строения разных отделов мозга и в первую очередь медуллярного уровня (рис. 27). В подобных случаях наблюдаются разные модификации внутреннего строения - от простого увеличения числа элементов до чрезвычайно сложных клеточных дифференциаций и четко выраженного ламинарного характера строения стволовых образований (рис. 28).
Естественно, что в состав продолговатого мозга входят и структуры, не относящиеся к сегментарным отделам, но включенные в различные системы мозга. Их удельный вес на уровне продолговатого мозга невелик, однако в ходе филогенеза происходит его неизбежное возрастание. Наконец, значительная часть продолговатого мозга составлена ретикулярной формацией (см. гл. 6).
66
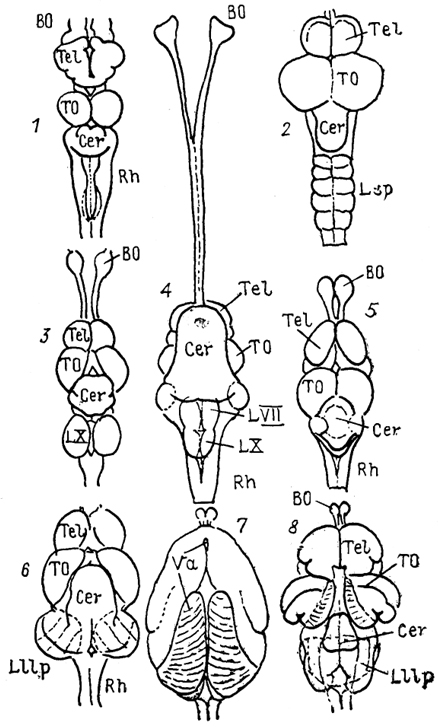
Рис. 27. Общий вид мозга разных представителей лучеперых рыб (Nieuwenhuys, Meek, 1985).
l -Amia, 2 - Trigla, 3 - Carassius, 4 - Clarlas, 5 - Xenomystus, 6 - Eigenmannia 7, 8 - Gnathonemus: до (7) и после (8)удаления заслонки.
Система черепномозговых нервов развивается как продолжение системы спинномозговых нервов, осуществляющих иннервацию туловища соответственно расположению миотомов.
Черепномозговые нервы в свою очередь формируются как бранхиальные, осуществляющие иннервацию области жаберных щелей, и эволюционируют параллельно с эволюцией жаберных дуг.
67
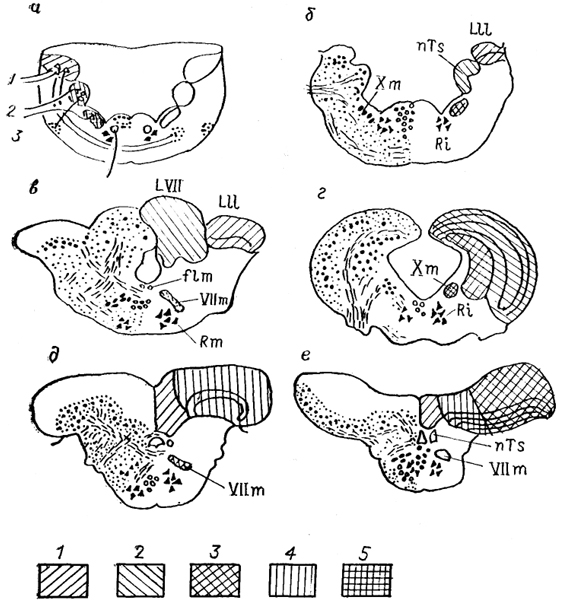
Рис. 28. Строение ромбовидного мозга разных групп рыб (Nieuwenhuys, Meek, 1985).
a - расположение колонн в мозге; 6-е - фронтальные сечения мозга Amia (б), Clarlas (в), Carassius (г), Xenomystus (д), Eigenmannla (e). 1-3 зоны продолговатого мозга; специальная соматосенсорная (1), висцеросенсорная (2), висцеромоторная (3); 4, 5 - электросенсорная область представительства ампуллярных (4) и бугорковых (5) рецепторов органов боковой линии электрических рыб.
В целом у ранних позвоночных четко выделяются бранхиальные нервы, гомологичные будущим тройничному (V), лицевому (VII), языкоглоточному (IX) и блуждающему (X). Начинающийся впоследствии процесс усложнения этой системы приводит к ее значительным преобразованиям. Так, путем слияния проекций чувствительных ветвей VII, IX и X пар черепномозговых нервов дифференцируется система вестибуло-кохлеарного (VIII) нерва, представленного уже в мозге круглоротых (рис. 29). Вероятно, позднее, судя по его слабой выраженности у бесчелюстных, формируется центральное представительство исключительно моторного отводящего (VI) нерва (рис. 30). У рептилий в качестве черепномозгового выделяется подъязычный (XII) нерв, представляющий собой объединение передних корешков трех ростральных сегментов спинного мозга. Отметим, что у некоторых рыб этот нерв уже присутствует в виде так Называемой задней доли, lobus posterior, но расположен не в краниальном, а в спинальном отделе (см, рис. 5, 6, 7).
68

Рис. 29. Общий вид
(А) и фронтальные сечения
(Б) мозга миноги
Ichthyomyzon unicuspis (Ronan, 1989).
1 - ядро заднего бугорка, 2 - субкомиссуральный орган, 3 - претекталъное ядро, 4 - ядро задней комиссуры, 5 - маутнеровская клетка; а-г - срезы в рострокаудальном направлении.
69
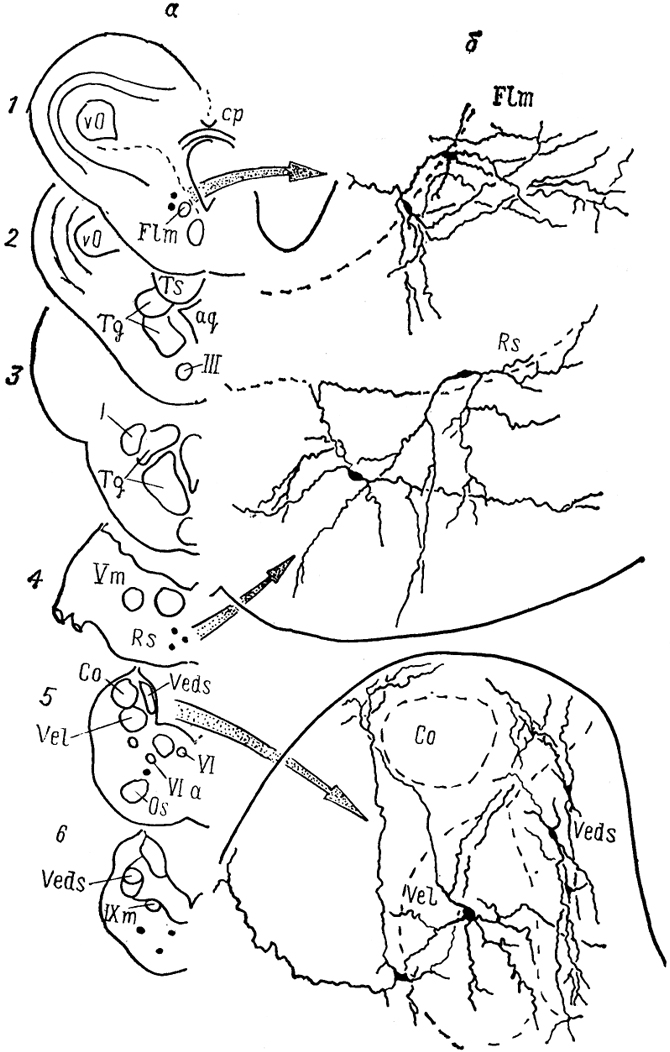
Рис, 30. Ядра
(а)и нейронный состав
(б)мозга лягушки
Rana esculenta (Toth e. a., 1985).
1-6- срезы в рострокаудальном направлении.
70
У бесхвостых амфибий местом его выхода является уровень II-III шейных сегментов. У рептилий, птиц и млекопитающих происходит вычленение добавочного (XI), чисто моторного нерва, как полагают, отделяющегося от блуждающего нерва.
Таким образом, в восходящем ряду позвоночных происходят процессы, приводящие к выделению и развитию имеющейся в настоящее время у высших системы черепномозговых нервов. Адаптивные изменения, обусловленные различием сред обитания, сказываются на всех компонентах этой системы, выражаясь не только в разной локализации выхода корешков из мозга, но и в разной степени развития их центральных проекций, а следовательно, в размерах (диаметре) нервов. На уровне продолговатого мозга такие различия особенно заметны в области производных крыловидной пластинки, где сосредоточены сенсорные ядра черепномозговых нервов.
71
