Все проводящие пути спинного мозга сосредоточены в белом веществе, разделяемом на три канатика: задний., боковой и передний. Основной объем канатиков занимают супраспи-нальные тракты, осуществляющие двустороннюю связь спинного
56
и головного мозга. Тракты, занимающие узкую полоску вокруг серого вещества, называются проприоспинальными (рис. 15).
Проприоспинальные тракты образованы аксонами интернейронов промежуточной зоны серого вещества. Распределение тел и окончаний проприоспинальных нейронов у позвоночных, начиная с самых ранних этапов эволюции, строго упорядочено. У высших позвоночных аксоны нейронов латеральных отделов пластин V-VII проходят в боковом канатике и оканчиваются в дорсолатеральных областях латеральных моторных ядер, ин-нервирующих мышцы-сгибатели. Нейроны центра пластины VII проецируются в вентромедкальную область этих ядер, связанную с мышцами-разгибателями. Интернейроны из медиальных районов пластин VII и VIII проецируются в медиальные моторные ядра, которые иннервируют осевую туловищную мускулатуру. При этом нейроны, образующие длинные восходящие и нисходящие проприоспинальные волокна, располагаются преимущественно в вентральных участках пластин V-VIII, а их аксоны оканчиваются в вентромедиальных районах пластин V-VII.
Проприоспинальные нейроны, относящиеся к разновидности ретикулярных изодендритных нейронов, создают морфологическую основу для широкой конвергенции на их телах и дендритах супраспинальных и сенсорных афферентов. Большинства проприоспинальных нейронов активируется супраспинальными трактами моносинаптически, а первичными афферентами - полисинаптически. Таким образом, появление и развитие в эволюции позвоночных системы проприоспинальных межсегментарных связей обеспечивает (наряду с другими механизмами) координацию сложных моторных реакций.
Супраспинальные тракты наибольшего развития и сложности достигают у высших млекопитающих - приматов, у которых описано их более двух десятков (рис. 15). Остановимся на наиболее важных из них.
Дорсальные канатики содержат восходящие тракты, волокна которых образованы аксонами сенсорных нейронов и коллатералями интернейронов пластин V и VI. Система дорсальных канатиков у высших позвоночных построена по соматотопическому принципу: чувствительные волокна от нижней части туловища и нижних конечностей проходят медиально, формируя тонкий пучок Голля (fasciculus gracilis), от верхней части тела и верхних конечностей - латерально, образуя клиновидный пучок Бурдаха (fasciculus cuneatus), причем в самих трактах пучки волокон от определенных участков тела и конечностей проходят также отдельно друг от друга. Функционально они тоже разделяются: волокна, несущие информацию от рецепторов волосяных фолликулов и конечностей, составляют поверхностную часть пучков, от мышечных рецепторов – среднюю
57
часть, а от рецепторов давления и вибрации - глубокую центральную часть. Оканчиваются волокна дорсальных канатиков в соответствующих ядрах продолговатого мозга.
В эволюции позвоночных происходит морфологическое обособление дорсальных канатиков от латеральных. Так, уже у хрящевых и некоторых костистых рыб они разделяются волокнами нисходящего бульбо-спинального тракта. Волокна дорсальных канатиков достигают продолговатого мозга и оканчиваются в зоне, которая по своей структурно-функциональной организации соответствует ядрам этого тракта у высших позвоночных. У наземных позвоночных уровень дифференцировки дорсальных канатиков тесно связан с развитием конечностей. Например, среди рептилий он наиболее, высок у крокодилов и наименее - у змей. Кроме того, начиная с амфибий, в организации этой системы все более отчетливо проявляется соматотопический принцип, Достигающий своего максимального выражения у млекопитающих и человека.
Система восходящих путей латеральных канатиков включает в себя ряд крупных трактов, соединяющих спинной мозг с продолговатым - спинно-ретикулярные, со средним - спинно-тектальные, с промежуточным мозгом - спинно-таламические, и мозжечком - спинно-мозжечковые. У низших позвоночных еще нет анатомического разделения трактов. У рептилий латеральные канатики уже дифференцированы на дорсо- и вентролатеральные пучки. Первый содержит в основном спинно-ретикулярные, спинно-тектальные и спинно-мозжечковые тракты, а второй - спинно-таламические и большую часть спинно-ретикулярных трактов.
Спинно-ретикулярные тракты обнаружены у всех изученных позвоночных - от круглоротых до человека. Они оканчиваются в каудальных отделах ретикулярной формации продолговатого мозга, моторных ядрах черепномозговых нервов, достигая в ряде случаев ретикулярной формации среднего мозга. Считается, что спинно-ретикулярные тракты позвоночных обеспечивают проведение информации от болевых рецепторов, а также пролриорецепторов.
Спинно-тектальные тракты описаны у большинства позвоночных, за исключением миног, лучеперых рыб и бесхвостых амфибий (возможно, что они утратили их в процессе эволюции независимо друг от друга). Спинно-тектальные тракты оканчиваются в центральном сером веществе мозга, в глубоких слоях тектума, тегментуме и интерколликулярном ядре.
Спинно-таламические тракты (спинно-таламический и спинно-ретикуло-таламический) обеспечивают болевую и температурную чувствительность и проведение информации от волосяных фолликулов, рецепторов давления и мышечных афферентов III группы. Спинно-ретикуло-таламичесюий тракт, как филогенетически наиболее древний, описан практически у всех
58
позвоночных, тогда как прямой спинно-таламический тракт обнаружен лишь у амниот и хрящевых рыб. Объем спинно-таламического тракта, а также локализация нейронов, образующих его, место окончания и количество передаваемой информации значительно варьируют даже у близкородственных групп позвоночных. Так, у млекопитающих проекции в специфические зоны таламуса (вентробазальный комплекс ядер) образованы нейронами пластин I и V спинного мозга, а в неспецифические зоны (медиальный комплекс ядер) -нейронами VII и VIII пластин. У птиц количество спинно-таламических нейронов очень невелико, полностью отсутствуют проекции из I пластины, при этом практически отсутствуют спинно-таламические проекции из шейных сегментов спинного мозга, иннервирующих крылья.
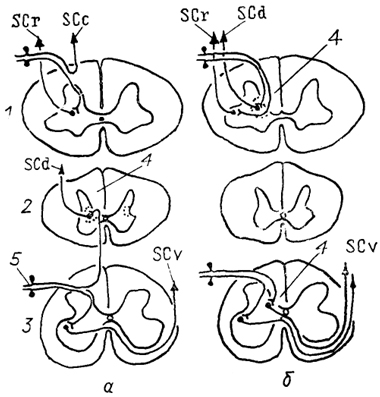
Рис. 23. Организация спинно-мозжечковых трактов у млекопитающих
(а) и птиц
(б) (Necker, 1989).
1-3 - шейный (1), грудной (2) и поясничный (3) отделы спинного мозга; 4 - ядро Кларка; 5 - чувствительные нейроны спинномозгового ганглия. Остальные обозначения, как на рис. 15.
Спинно-мозжечковые тракты есть у всех позвоночных, за исключением миног и миксин. У низших позвоночных спинно-мозжечковые волокна вместе с другими восходящими трактами входят в состав латерального канатика белого вещества спинного мозга и отделяются от них только в продолговатом мозге на уровне ядра V нерва. Разделение на вентральный и дорсальный компоненты намечается уже у хрящевых рыб и завершается у млекопитающих у которых выделяют дорсальный, вентральный и ростральный спинно-мозжечковые тракты и спинно-кунео-мозжечковый тракт (рис. 23). Дорсальный тракт начинается от крупных нейронов ядра Кларка в пластине IV серого вещества, оканчивается в I-IV и частично в V дольках мозжечка и несет информацию от отдельных мышц туловища и нижних конечностей. Вентральный образован нейронами нижних сегментов спинного мозга, расположенными в пластине V, и связан с проведением информации от группы мышц. Ростральный спинно-мозжечковый и спинно-кунео-мозжечковый тракты получают информацию от рецепторов верхних конечностей
59
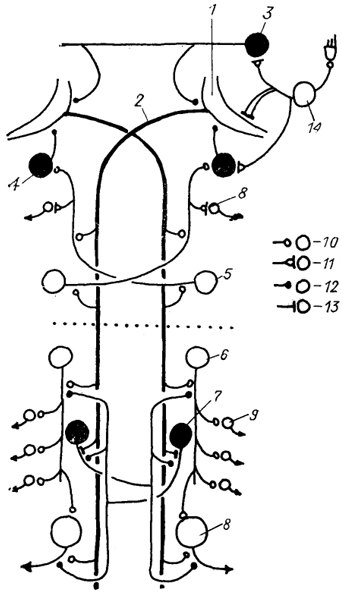
Рис. 24. Схема нейронной сети маутнеровских нейронов в спинном мозге костистых рыб (Faber e. а., 1989).
1 - маутнеровская клетка и ее аксон(2); 3-5 - комиссуральные (3) ,коллатеральные (4) и краниальные (5) интернейроны продолговатого мозга; 6, 7 - нисходящие (6) и реципрокные (7) интернейроны спинномозгового сегмента; 8, 9 - первичные (8) и вторичные (5) мотонейроны; 10-13 - синаптические контакты химического (10), электрохимического (11), химического тормозного (12) и электротонического (13) типов; 14 - нейроны чувствительного ганглия вестибулярного нерва. Темные клетки - тормозные, светлые - возбуждающие. Пунктиром обозначена граница головного и спинного мозга.
и начинаются в шейных сегментах спинного мозга. Все тракты и особенно дорсальный имеют четкую соматотопическую организацию. У птиц организация спинно-мозжечковых трактов иная, чем у рептилий и млекопитающих (рис. 23, б).
Нисходящие супраспинальные тракты опосредуют влияние головного мозга на моторные центры спинного мозга, и их активность связана прежде всего с такими рефлекторными актами, как плавание, бег, шагание, прыжки, полет. Система нисходящих трактов сформировалась на ранних этапах эволюции позвоночных и имеет единый план строения, включая несколько основных трактов: ретикуло-, вестибуло-, текто- и рубро-спинальные.
У низших позвоночных основную массу нисходящих трактов составляют ретикуло-спинальные тракты. Среди нейронов, образующих эти тракты, особо выделяются мюллеровские и маутнеровские клетки, расположенные в стволе мозга и обладающие длинными и нисходящими аксонами, которые моносинаптически связаны с мотонейронами спинного мозга, управляющими движением туловища и хвоста. У костистых рыб аксон каждой маутнеровской клетки ипсилатерально образует химические возбуждающие синапсы на спинальных нейронах двух типов: первичных мотонейронах и так называемых нисходящих интернейронах, аксоны которых на протяжении двух и более сегментов контактируют с первичными и вторичными мотонейронами (рис. 24).
Анализ структуры ретикуло-спинальных трактов высших позвоночных
60
показал, что данная система является наиболее консервативной. Это подтверждается ранним формированием ретикуло-спинальных трактов в онтогенезе.
Вестибуло-спинальные тракты развиты практически у всех позвоночных и разделяются на несколько пучков. У млекопитающих описаны прямой тракт (из латерального вестибулярного ядра) и два перекрещенных (из латерального и нисходящего вестибулярных ядер). Стимуляция этих ядер оказывает билатеральное действие на нейроны спинного мозга, связанные с движением головы, плечевого пояса и верхних конечностей. Тракт заканчивается на мотонейронах или интернейронах пластин VI-VIII.
Текто-спинальные тракты берут начало из различных отделов tectum opticum и опосредуют его влияние на мотонейроны спинного мозга. У млекопитающих они частично перекрещены. Обширные текто-спинальные тракты описаны у хрящевых и двоякодышащих рыб, амфибий (хвостатых и бесхвостых) и рептилий. По-видимому, они отсутствуют у некоторых костистых рыб (например, у лососевых). У кругло>ротых (по крайней мере, у миног) текто-спинальные волокна достигают верхних сегментов спинного мозга.
Перекрещенный рубро-спинальный тракт, сформированный нейронами красного ядра, обнаружен у бесхвостых и хвостатых амфибий, рептилий, птиц и млекопитающих; он, вероятно, отсутствует у безногих амфибий, змей и рыб. Прямой рубро-спинальный тракт описан у некоторых хрящевых (например, у скатов) и костных рыб, которые используют свои парные грудные придатки для передвижения. Таким образом, рубро-спинальные тракты эволюционно связаны с появлением конечностей и поясов конечностей, и степень их развития коррелирует со степенью развития мозжечка (однако присутствие рубро-спинального тракта у змеи Nerodia fasciata опровергает мнение о том, что он может быть связан только с регуляцией движений конечностей). Заканчиваются тракты в спинном мозге на интернейронах пластин V-VII.
Кортико-спинальный (пирамидный) тракт появляется у птиц и млекопитающих и является наиболее молодой из всех провОДЯЩИХ систем спинного мозга. Размер тракта увеличивается в ряду млекопитающих, тесно коррелируя с развитием моторной активности вида и со способностью к тонким дифференцированным движениям конечностей, и достигает максимума у человека. В спинном мозге зона прямых проекций тракта значительно варьирует у разных видов (рис. 25). У низших млекопитающих (например, у сумчатых) кортико-спинальные волокна достигают только шейных и грудных сегментов, проходя в дорсальных канатиках, и оканчиваются в задних рогах спинного мозга. У высших млекопитающих кортико-спинальный
61
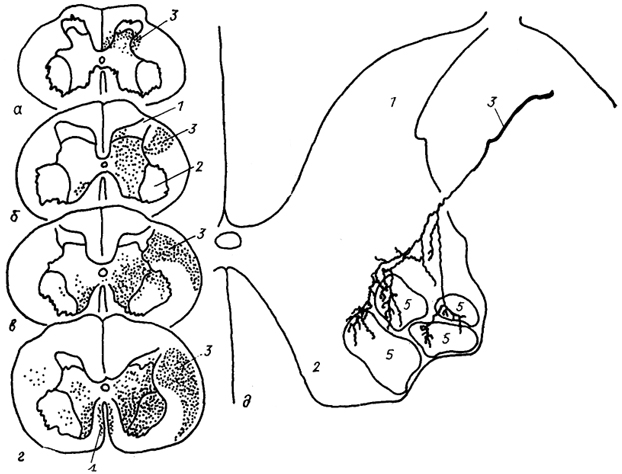
Рис. 25. Окончание кортико-спинального тракта в шейных сегментах спинного мозга опоссума
(а), кошки
(б), макаки
(в), шимпанзе
(г) и характер ветвления одного волокна в моторных ядрах мозга макаки
(д) (Shinoda е. а., 1979; Kuypers, 1981).
1, 2 - задние (1) и передние (2) рога спинного мозга; 3, 4 - латеральный (3) и передний (4) кортико-апинальные тракты; 5 - моторные ядра.
тракт проходит вдоль всего спинного мозга, разделяясь на два! компонента (латеральный и вентральный).
Кортико-спинальные волокна начинаются в разных областях новой коры, и, таким образом, пирамидный тракт по существу представляет собой комплекс отдельных путей, каждый из которых отличается по характеру окончаний в спинном мозге. Так, у высших приматов волокна от моторных зон коры проецируются в промежуточную зону и моторные ядра, волокна от соматосенсорных зон - в пластины I-IV. Дифференцированное распределение кортико-спинальных проекций свидетельствует об их влиянии на разные спинальные механизмы, вплоть до регуляции сенсорных афферентов в пластинах I и П. Более того, волокна из моторных зон коры оканчиваются преимущественно на телах и проксимальных дендритах мотонейронов, тогда как проекции из соматосенсорных полей - на дистальных дендритах. У большинства млекопитающих пирамидный тракт созревает в постнатальный период, причем в процессе его развития происходит элиминация многих кортико-спинальных
62
волокон, выходящих из некоторых корковых зон-(например, зрительной коры).
Таким образом, уже в строении спинного мозга различных позвоночных отчетливо прослеживаются тенденции последовательной дифференциации его элементов и связей, а также усложнение межнейронных взаимодействий, обусловленные системным повышением уровня организации ЦНС.
63
