Спинной мозг, Medulla spinalis, является центром, воспринимающим разнообразную соматическую информацию из внешней и внутренней среды и передающим ее в вышележащие отделы ЦНС. В нем сосредоточены моторные (эфферентные) центры, управляющие рефлекторной деятельностью поперечнополосатых мышц туловища и конечностей, и ассоциативные центры симпатического и парасимпатического отделов вегетативной нервной системы.
Анатомически спинной мозг разделяется на отделы, количество сегментов в которых соответствует числу позвонков у представителей данной группы позвоночных. Так, у птиц выделяют 12 шейных, 8 грудных, 12 поясничных и 6 копчиковых сегментов; у приматов - 8 шейных, 12 грудных, по 5 поясничных и крестцовых и 2-3 копчиковых, а у некоторых рептилий (змей) число спинномозговых сегментов может достигать 500.
У большинства позвоночных (рыбы, хвостатые амфибии, рептилии, птицы) спинной мозг занимает весь позвоночный канал, тогда как у бесхвостых амфибий и млекопитающих он занимает только часть канала из-за неравномерности роста спинного мозга и позвоночника в период эмбрионального развития. Так, у большинства млекопитающих спинной мозг оканчивается на уровне поясничных позвонков, однако у примитивных млекопитающих (нааример, ехидна) конец спинного мозга находится на уровне грудного отдела позвоночника. Наиболее короткий спинной мозг отмечен у южноамериканской лягушки Pipa pipa - он оканчивается на уровне 3-го позвонка. При этом для всех позвоночных выполняется одно правило: спинномозговые нервы каждого сегмента выходят из позвоночного канала через межпозвонковые отверстия на уровне
42
"своих" позвонков, в связи с чем в каудальной части позвоночного канала формируется особая анатомическая структура - пучок спинномозговых корешков, конский хвост.
На уровне каждого сегмента с обеих сторон спинного мозга отходит пара спинно-мозговых корешков: задний (дорсальный) и передний (вентральный) (рис. 15). У круглоротых дорсальные и вентральные корешки спинного мозга отходят асимметрично, тогда как у всех других позвоночных - симметрично.
В районе межпозвонковых отверстий спинномозговые корешки у большинства позвоночных (за исключением бесчерепных, миног и некоторых рыб) соединяются в единый спинномозговой нерв.
В составе задних корешков спинного мозга находятся спинномозговые ганглии (каждый корешок имеет по одному ганглию), содержащие чувствительные (афферентные) нейроны, которые воcпринимают
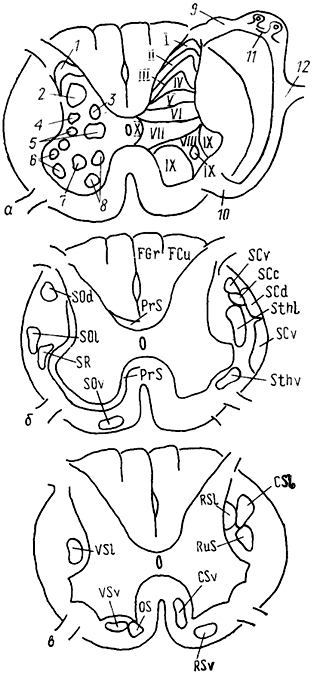
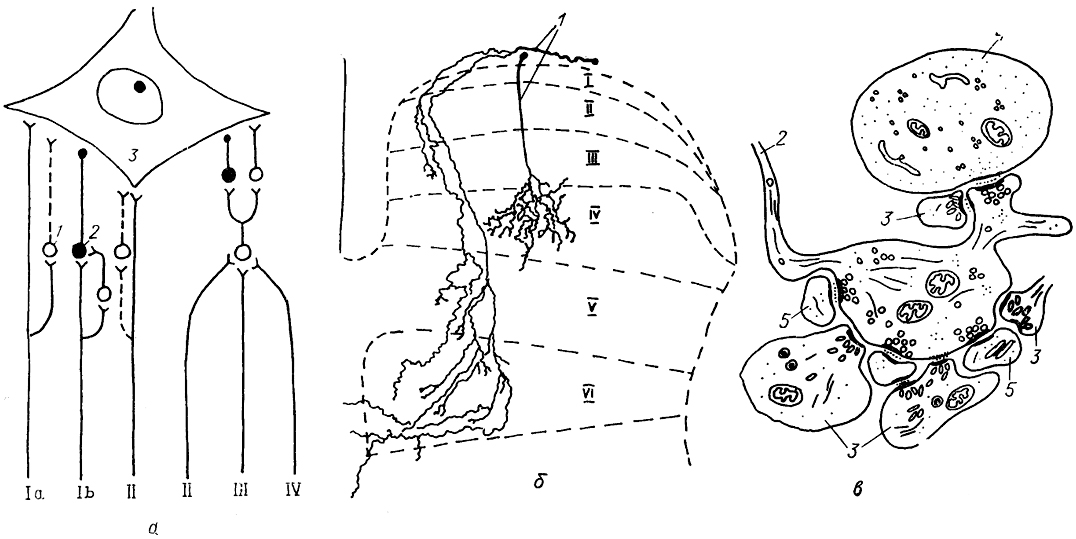
Рис. 15. Схема организации серого вещества
(а) и топография восходящих
(б) и нисходящих
(в) трактов спинного мозга.
а: 1-Х - пластины серого вещества по классификации Б. Рекседа; 1-4 - желатинозная субстанция (1), собственное ядро (2), ядро Кларка (3), ретикулярное (4) ядро задних рогов; 5 - промежуточная зона, 6-8 - моторные ядра передних рогов: латеральные (6) , центральные (7), медиальные (8); 9, 10 - задний (9) и передний (10) спинномозговые корешки; 11 - спинальный ганглий, 12 - спинномозговой нерв.
б: восходящие тракты - спинно-мозжечковые: дорсальный SCd, вентральный SCv, ростральный SCr и кунеальный SCc; спинно-таламические: латеральный SThI и вентральный SThv. спинно-ретикуло-таламический SRT, спинно-тектальный ST; спинно-оливарные: дорсальный SOd, латеральный SOl и вентральный SOv, спинно-ретикулярный SR, нежный FGr и клиновидный FCu пучки; проприоспинальные PrS (собственные) пути спинного мозга (восходящие и нисходящие).
нисходящие тракты - кортико-спинальные: латеральный CSl и вентральный CSv, руброспинальный RuS; вестибуло-спинальные: латеральный VSI и передний VSv; ретикуло-спинальные: латеральный RSl и вентральный RSv; оливо-спинальный OS.
43
разнообразную информацию из кожи, опорно-двигательного аппарата, внутренних органов. Спинальные ганглии развиваются из элементов ганглионарной пластинки. Принципиальным является тот факт, что у ряда низших позвоночных (взрослые круглоротые, личинки рыб, амфибий) чувствительные нервные клетки располагаются не только в спинальных ганглиях, но и внутри спинного мозга. Эти клетки, получившие название "клетки Рогон-Берда", или дорсальные чувствительные клетки, находятся в дорсолатеральной части спинного мозга и представляют собой крупные (8-20 мкм) нейроны. От их тел отходят по два отростка, которые направляются вдоль спинного мозга, образуя синапсы (в основном
Таблица 3. Классификация афферентных волокон спинного мозга
| Тип |
Диаметр, мкм |
Миелинизация |
Функциональная характеристика |
| по Гассеру |
по Ллойду |
| Аα |
Ia, Ib |
12 - 20 |
+ |
Проведение возбуждения от: первичных рецепторов мышц и сухожилий; |
| Аβ |
II |
6 - 12 |
+ |
от вторичных рецепторов мышц и рецепторов кожи; |
| Аδ |
III |
4 - 6 |
+ |
от рецепторов кожи и внутренних органов; |
| С |
IV |
1,5 - 4 |
- |
от рецепторов сосудов, кожи, мышц |
электротонические) на телах и отростках спинальных интернейронов. Последние в свою очередь контактируют с мотонейронами, таким образом формируя первичную рефлекторную дугу сомато-моторного рефлекса. Электрофизиологические исследования показали, что клетки Рогон-Берда и замещающие их на более поздних этапах эмбриогенеза так называемые дорсальные чувствительные клетки связаны с механорецепцией,. тогда как чувствительные клетки в спинномозговых ганглиях воспринимают болевые и температурные стимулы. На теле клеток Рогон-Берда обнаружены многочисленные синаптические контакты неизвестного происхождения. Полагают, что они модулируют выделение медиатора (вещество Р) из клеток. У высших позвоночных на телах чувствительных нейронов в; спинномозговых ганглиях обнаружены синаптические контакты, которые образованы аксонами спинальных нейронов, приходящими по обоим корешкам.
Структура спинномозгового ганглия достаточно сложная. Морфологически в нем выделяются три основные группы нейронов: крупные и мелкие псевдоуниполярные и атипичные. Последние осуществляют внутриганглионарные связи. Более многочисленные
44
(85%) псевдоуниполярные нейроны являются сенсорными и обеспечивают поступление в спинной мозг информации об активации рецепторов разных типов (табл. 3).
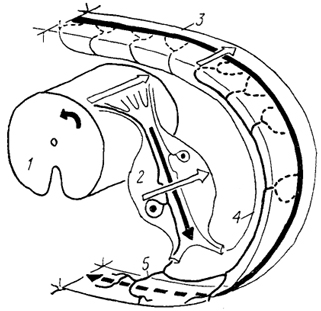
Рис. 16. Схема организации спинномозгового ганглия кошки (Kausz, Rethelyi, 1985).
1 - спинной мозг; 2 - ганглий; 3 - стенка туловища; 4, 5 - дорсальная (4) и вентральная (5) ветви чувствительного нерва. Стрелки показывают соответствие зон иннервации положению клеток в ганглии и спинном мозге.
Расположение клеток, связанных с разными видами чувствительности и иннервирующих определенные участки тела, строго упорядочено. Так, у кошки дорсовентральный градиент распределения зон иннервации в коже соответствует проксимальнодистально-му градиенту распределения клеток в ганглии, а каудальноростральный градиент зон иннервации - медиолатеральному градиенту распределения клеток (рис. 16).
На сегментарном уровне организации спинного мозга сохраняется закономерный характер проекций. Во-первых, на уровне каждого сегмента по афферентным волокнам всех типов в спинной мозг поступает информация от строго определенного участка тела. Во-вторых, волокна, иннервирующие более дистально расположенные участки тела имеют намного меньшие рецептивные поля, чем проксимальные. Этим обеспечивается более высокая точность различения сигналов от дистальных участков конечностей (пальцы), связанных, как правило, с выполнением более тонких и дифференцированных движений, чем от проксимальных отделов конечностей.
Наиболее подробно у позвоночных изучено взаимодействие первичных афферентных волокон с мотонейронами спинного мозга. У круглоротых, как указывалось выше, часть клеток Рогон-Берда и клеток спинальных ганглиев не только устанавливает контакты с интернейронами, но и активирует мотонейроны посредством образования аксо-соматичеоких синапсов электро-химического типа.
45

Рис. 17. Окончание афферентных волокон в спинном мозге карпа
(а) и на" одиночном мотонейроне спинного мозга лягушки
(б) (Моторина, 1982; 1983).
1 - афферентные волокна и их ветви; 2 - мотонейроны; 3,4 - задние (3) и передние (4) рога серого вещества; 5, 6 - аксон (5) и дендрит (6) мотонейрона.
46
У рыб выявлены афферентные волокна двух типов. Одни из них оканчиваются на интернейронах медиальной части дорсального рога серого вещества спинного мозга, другие образуют аксо-соматические и аксо-дендритные контакты смешанного типа на мотонейронах вентрального рога. Часть афферентных волокон может переходить на противоположную сторону спинного мозга (рис. 17). У ряда изученных хрящевых рыб (например, некоторые скаты из группы Rajaformes и акулы Squalus) обнаружены прямые моносинаптические связи с мотонейронами, тогда как у других (например, акулы Scyliorhinus canicula) таких связей не выявлено.
У амфибий афферентные волокна образуют контакты не только с дистальными дендритами мотонейронов (как у большинства рыб), но и с их проксимальными ветвями и телами. Число контактов отдельного волокна с мотонейроном может достигать 60 - 70 (рис. 17). Показано, что афферентные волокна от определенной мышцы ветвятся среди большой группы мотонейронов, связанных с иннервацией различных мышц, но образуют контакты только с мотонейронами "своей" мышцы. У амфибий в спинном мозге, впервые среди позвоночных, намечается четкое топографическое разделение зон ветвления афферентов разного типа. Первичные афференты мышечных волокон (тип А) контактируют с мотонейронамн в вентральном. роге, тогда как афференты кожной и мышечной чувствительности (типы Iа, II) связаны с мотонейронами через систему интернейронов, и зона их ветвления ограничена дорсальным рогом.
Распределение афферентов в спинном мозге рептилий сходно с таковым у амфибий. Часть волокон оканчивается в дорсальном роге, другие достигают вентральных рогов. Электро-физиологически показано наличие прямых моносинаптических связей афферентов с мотонейронами у ящериц, крокодилов и черепах.
У млекопитающих и птиц происходит дальнейшая дифференциация входов. Основная масса проекций от кожных рецепторов (связанных с ноци-, механо- и терморецепцией) направляется в ядра дорсального рога серого вещества, а проекции таких специализированных рецепторов, как сухожильные, расположены в основании дорсального рога. Таким образом, большинство афферентов не достигает вентральных рогов спинного мозга, их контакт с мотонейронами осуществляется полисинаптически, через систему интернейронов (рис. 18). Ветвления отдельных афферентных волокон строго упорядочены и формируют сложную систему пространственных модулей, обеспечивающих высокий уровень специфичности межнейронных связей.
Серое вещество спинного мозга состоит из тел и отростков нейронов и у всех позвоночных расположено вокруг центрального спинномозгового канала, являющегося остатком полости
47
48
нервной трубки. Основная эволюционная тенденция в развитии серого вещества состоит, во-первых, в миграции клеток от центрального канала на периферию и, во-вторых, в прогрессивной дифференциации клеточных популяций на отдельные зоны и ядра. Так, у бесчерепных все клетки спинного мозга располагаются вокруг центрального канала, тогда как у круглоротых часть нейронов активно мигрирует в толщу спинного мозга, формируя две латерально расположенные группы. У остальных позвоночных в спинном мозге образуются отдельные популяции нейронов дорсальных и вентральных рогов. Позднее, у наземных позвоночных, в ряде отделов спинного мозга появляются боковые рога.
В 1952-1954 гг. Б. Рексед предложил новую универсальную классификацию серого вещества спинного мозга, разделив его на десять пластин (I-X), различающихся по структуре составляющих их элементов и по связям (рис. 15, а).
Пластина I представлена узкой полоской нейронов с характерными тангенциально ориентированными дендритами. На интернейронах пластины I оканчиваются афференты типов Аβ, Аδ и С. Аксоны интернейронов образуют восходящие тракты спинного мозга, а также посылают коллатерали в более глубокие слои спинного мозга (рис. 19). Помимо сенсорных волокон в I пластине обнаружены многочисленные афференты от вышележащих отделов спинного и головного мозга, в том числе от структур лимбической системы, через которые осуществляется контроль болевых ощущений.
Пластины II и III построены из нейронов среднего размера, дендриты которых могут проникать как в зону пластины I, так и в более глубокие слои серого вещества. Аксоны этих клеток распространяются на большие расстояния и захватывают несколько сегментов, образуя многочисленные внутри- и межсегментные связи. Часть аксонов образует систему восходящих трактов.
Обилие входящих в пластины I-III афферентных волокон, разнообразие нейронных типов, наличие синоптических комплексов конвергентного и дивергентного типов и модуляторных контактов определяют сложный характер межнейронных взаимодействий. Так, в вентральной части пластины III особенно многочисленны специфические формы аксо-аксональных синапсов между нейронами и афферентами, а также синапсы дендро-дендритного типа (рис. 18). Таким образом, значение нейронных комплексов пластин I-III состоит в контроле поступающей в спинной мозг сенсорной информации различного вида, в ее переработке и передаче в спинной и головной мозг.
Пластины IV-VI представлены скоплениями нейронов мультиполярного типа, дендриты которых ориентированы преимущественно в дорсальном направлении и проникают в зону пластин I-III. На телах и дендритах нейронов оканчиваются
49
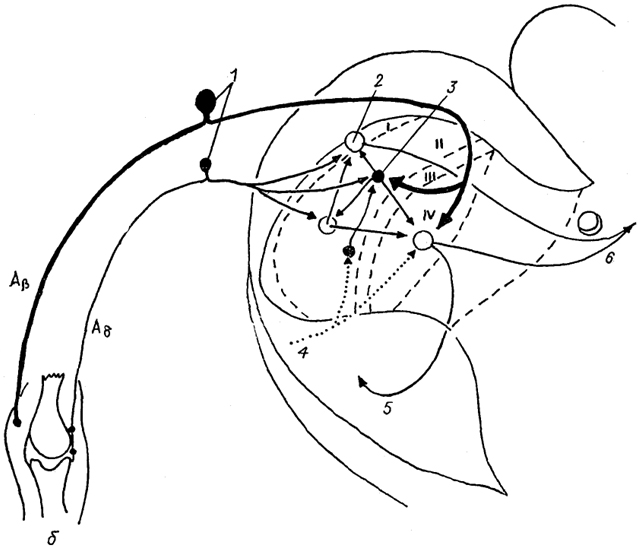
Рис. 19. Межнейронные связи в I-IV пластинах заднего рога спинного мозга (Delwaide, Schoenen, 1989).
Aβ, Aδ - афференты; 1 - чувствительные нейроны спинального ганглия; 2, 3 - возбуждающие (2) и тормозные (3) интернейроны; 4 - нисходящий бульбо-спинальный тракт; 5, 6 - восходящие спинно-ретикуло-таламический (5) и спинно-таламический (6) тракты.
многочисленные терминали первичных афферентов из групп Аα, Аβ, Аδ, аксоны интернейронов собственного и соседних сегментов, а также волокна нисходящих трактов. Окончания афферентов строго упорядочены: в вентральный отдел пластины V проецируются болевые и высокопороговые механорецепторы, в средний - рецепторы кожи, в дорсальный - низкопороговые: механорецепторы волосяных фолликулов. Эти отделы спинного мозга являются источником основной массы проприоспинальных (собственных) путей спинного мозга, образуя три системы связей: короткие - в пределах 1-2 сегментов, промежуточные- 4-12 сегментов, и Длинные. Последние представлены аксонами, идущими в составе тонкого пучка Голля в продолговатый мозг, в то время как их коллатерали достигают промежуточного мозга. Нейроны, аксоны которых образуют систему длинных связей, локализованы в основном в пластине IV шейных и пластинах V и VI пояснично-крестцовых сегментов спинного
50
мозга. Промежуточные и короткие проприоспинальные, пути связаны с проведением информации от рецепторов мышц, суставов, сухожилий. Часть волокон оканчивается на крупных нейронах ядра Кларка в пластине V, образующих в свою очередь крупный восходящий дорсальный спинно-мозжечковый тракт. Нейроны ядра Кларка получают проекции от афферентов флексорного рефлекса (FRA), связанных с иннервацией конечностей, причем разные афференты проецируются на различные клетки этого ядра.
Пластины VII и VIII, как полагают, не получают прямых афферентных проекций и занимают промежуточное положение между задними и передними рогами серого вещества спинного мозга. Здесь широко представлены терминальные ветвления проприо- и супраспинальных трактов. Аксоны интернейронов VII и VIII пластин оканчиваются на мотонейронах и образуют восходящие и нисходящие тракты различного типа. Одной из разновидностей этих интернейронов являются клетки Реншоу, получающие коллатерали аксонов мотонейронов и осуществляющие обратное торможение последних. Латеральные отделы пластины VII на уровне грудных - верхних поясничных сегментов образуют боковые рога и состоят из скоплений ассоциативных преганглионарных нейронов вегетативной нервной системы. Современные исследования, проведенные с помощью внутриклеточных инъекций пероксидазы хрена, показали сложную структурно-функциональную организацию боковых рогов. Их нейроны относятся к ретикулярному типу, для которого характерно широкое распространение дендритав, достигающих даже пластины I задних рогов. На дендритах и телах преганглионарных нейронов обнаружены многочисленные контакты различного происхождения. В целом подробный анализ нейронной структуры промежуточной зоны спинного мозга показал, что это образование ретикулярного типа, переходящее непосредственно в ретикулярную формацию ствола головного1 мозга и являющееся связующим звеном между сенсорными формациями заднего рога и моторными ядрами переднего рога.
Вентральные рога спинного мозга представляют особый интерес, так как здесь располагаются группы спинальных мотонейронов, управляющих деятельностью поперечнополосатой скелетной мускулатуры. В спинном мозге млекопитающих мотонейроны занимают район пластин VIII и IX и дифференцированы на группы ядер: медиальную, центральную и латеральную (рис. 15, а). Мотонейроны являются наиболее крупными клетками спинного мозга и разделяются на три группы: амотонейроны, крупные и обеспечивающие быстрые сокращения мышц; р-мотонейроны, мелкие и обеспечивающие медленные сокращения мышц, и у-мотонейроны, связанные с иннервацией мышечных веретен. На примере организации моторных ядер наиболее отчетливо проявляется принцип соматототщи: каждая
51
мышца или группа мышц иннервируется определенной группой мотонейронов (мотонейронный пул). В пределах сегмента организация мотонейронных пулов млекопитающих подчиняется так называемому правилу Романеса: нейроны, иннервирующие дистальные мышечные группы конечностей, располагаются в спинном мозге дорсально по отношению к нейронам, иннервирующим проксимальные мышечные группы. При этом мотонейроны мышц-сгибателей и -разгибателей, часто относящихся к одной мышечной группе, в структуре моторных ядер также располагаются изолированно друг от друга.
Пластина X - центральное серое вещество спинного мозга занимает небольшой участок вокруг центрального канала. Здесь располагаются мелкие интернейроны, связывающие ее с другими участками серого вещества, а также проходят пучки комиссуральных волокон, соединяющих правую и левую половины спинного мозга.
У круглоротых в спинном мозге намечается дифференциация мотонейронов на популяции, иннервирующие отдельные миомеры. Каждая такая популяция включает в себя группы первичных (крупных) и вторичных (мелких) мотонейронов, иннервирующих внутри отдельного миомера различные зоны (дорсальную или вентральную) и типы мышц. Первичные и вторичные мотонейроны образуют множественные контакты (от 3 до 8) на волокнах быстрых "белых" мышц - полинейронная иннервация. Мононейронная иннервация, осуществляемая только первичными мотонейронами, характерна для медленных "красных" мышц. Однако у круглоротых нет обособления отдельных популяций мотонейроров, иннервирующих разные типы мышц в пределах сегмента.
У хрящевых рыб дифференциация белого и серого вещества в спинном мозге нечеткая (рис. 20). В вентральных рогах имеются три группы клеток: дорсомедиальная, вентролатеральная и центральная. Для мотонейронов характерна большая вариабельность формы и размеров, а также тенденция к тесному расположению их тел и отростков. Особенность мотонейронов рыб состоит в том, что они увеличиваются в размерах в течение всей жизни организма (в отличие от нейронов других позвоночных).
У акул и скатов мотонейроны, иннервирующие красные и "белые мышцы миомеров, уже пространственно разграничены (рис. 20, в, г). Красные мышцы иннервируются мелкими мотонейронами, сосредоточенными в латеральной части мотонейронного пула, белые - крупными мотонейронами, занимающими медиальные районы. Более того, аксоны мотонейронов, иннервирующие
52
разные участки миомеров, имеют разный диаметр и проходят в разных участках вентральных корешков.
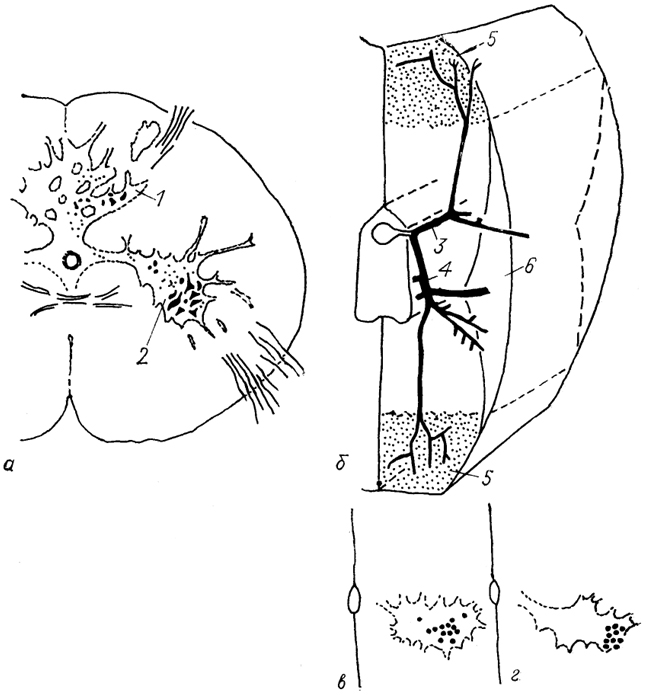
Рис. 20. Цитоархитектоника спинного мозга
(а) , ветвление вентрального корешка спинномозгового нерва
(б) и локализация мотонейронов, иннервирующих "красные"
(в) и "белые"
(г) мышцы у акулы (Mos, Willamson, 1986).
1, 2 - задние (1) и передние (2) рога серого вещества; 3, 4 - дорсальная (3) и вентральная (4) ветви вентрального корешка спинномозгового нерва; 5, 6 - зона "белых" (5) и "красных" (6) мышц миотома.
Мотонейроны костистых рыб расположены в вентральных рогах серого вещества спинного мозга и представлены двумя популяциями. Первую популяцию составляют крупные первичные мотонейроны, локализованные исключительно в дорсальной части мотонейронной колонны. Их сильно развитые дендриты проникают в белое вещество и часто достигают противоположной стороны спинного мозга. Таких мотонейронов немного: например, у золотой рыбки на 1/2 сегмента приходится 8-12
53

Рис. 21. Нейронная структура моторных ядер шейных сегментов спинного мозга хвостатых амфибий из группы саламандр:
Rhyacotriton olympicus (a), Ptethodon jordani
(б), Batrachoseps attenuatus
(в) (Toth e. a., 1985).
1, 2 - медиальная (1) и латеральная (2) группы мотонейронов; 3 - дорсальный корешок; 4 - спинномозговой канал. Точками обозначена зона серого вещества.
клеток. Они образуют аксо-аксональные контакты электротонического или смешанного типа с маутнеровскими нейронами. Вторая популяция представлена мелкими вторичными мотонейронами, занимающими в мотонейронной колонне сегмента вентральное положение. Отдельные первичные мотонейроны иннервируют определенные участки миомеров, а разные популяции мотонейронов иннервируют в миоме-ре различные типы мышц: первичные - белые, а вторичные - и белые и красные мышцы. Отдельное мышечное волокно иннервируется несколькими мотонейронами - полинейронная иннервация. Электрические органы, имеющиеся у некоторых рыб (B частности, у электрических скатов, сомиков) и развивающиеся из производных осевой мускулатуры, иннервируются специальной популяцией мотонейронов, расположенных в центральной зоне спинного мозга и связанных друг с другом многочисленными электротоническими контактами сомато - соматического и дендро-дендритного типов.
У амфибий, как и
54
у других низших позвоночных, мотонейроны разделяются на два основных класса: первичные (крупные) мотонейроны, дендриты которых распространяются на большие расстояния, захватывая несколько сегментов и переходя на противоположную сторону спинного мозга, и вторичные (мелкие) мотонейроны, аксоны которых не контактируют с маутнеровскими волокнами.
У хвостатых амфибий популяции мотонейронов формируют две группы, или колонны: вентромедиальную и вентролатеральную. Первая содержит в основном более мелкие грушевидные клетки, а вторая представлена крупными веретеновидными нейронами, причем степень развития той или иной мотонейронной колонны сильно варьирует даже у представителей одной труппы животных (рис. 21). Иннервация мышц у хвостатых амфибий полинейронная, и популяции мотонейронов содержат как первичные, так и вторичные мотонейроны.

Рис. 22. Нейронная структура вентромедиального (а) и вентролатерального
(б) мотонейронного ядра спинного мозга ящерицы (Cruce, 1979).
1, 2 -задние (1) и передние (2) рога спинного мозга; 3 - медиальные проприоспинальные тракты; 4 - спинномозговой канал; VII-IX -пластины Рекседа.
У бесхвостых амфибий в спинном мозге намечается пространственное разделение мотонейронных популяций, иннервирующих разные группы мышц. Клетки, иннервирующие осевую туловищную мускулатуру, расположены в мотонейронной колонне сегмента вентральнее и медиальнее мотонейронов, иннервирующих конечности, хотя внутри пула еще нет морфологической дифференцировки нейронов, иннервирующих отдельные мышцы. Мотонейроны образуют многочисленные электротонические, химические и смешанные контакты с клетками соседних сегментов и противоположной стороны спинного мозга, создавая основу для координированной деятельности мышц туловища и конечностей.
Мотонейроны спинного мозга рептилий организованы в две
55
моторные группы: вентромедкальную и вентролатеральную, нейроны которых отличаются друг от друга по размеру и характеру ветвления дендритов (рис. 22). Мотонейроны первой группы иннервируют осевую мускулатуру, второй - мускулатуру конечностей. У рептилий мотонейроны, иннервирующие отдельные мышцы, уже расположены раздельно. Однако в связи с тем, что у них самая сложная среди позвоночных мышечная система, имеется множество вариантов моторной иннервации. Так, красные медленные мышечные волокна рептилий иннервируются полинейронально и вдоль всей поверхности волокна, тогда как белые фазные волокна, разделяемые на быстрые и медленные, иннервируются мононейронально и локально. Очень часто в отдельных мышцах красные и белые волокна перемешаны друг с другом и иннервируются разными мотонейронами. Считается, что такая иннервация мышц у рептилий обеспечивает синхронизацию их работы (например, при движении туловища у змей). Подчеркнем, что у рептилий впервые среди позвоночных дендриты большинства мотонейронов не проникают на противоположную сторону спинного мозга, и это позволяет осуществлять более тонкую регуляцию деятельности мотонейронов спинномозгового сегмента через систему интернейронов.
Организация мотонейронных популяций, иннервирующих осевую мускулатуру млекопитающих, подробно изучена у крыс и кошек. Мотонейроны, связанные с определенными мышцами, занимают строго определенное положение в мотонейршном пуле. При этом мышцы, которые развиваются в соседних участках миомеров, иннервируются рядом расположенными нейронами, несмотря на возможное изменение положения данных мышц в туловище взрослого животного. У млекопитающих γ-мотонейроны, связанные с интрафузальными мышечными волокнами, рассеяны среди других нейронов данного пула. Интересно, что часть осевой мускулатуры млекопитающих - шейные мышцы - сохраняет филогенетически древний принцип иннервации - двумя разными типами мотонейронов, расположенных в разных ядрах. Остальная, как быстрая белая, так и медленная красная мускулатура не имеет двойной иннервации. Таким образом, в процессе эволюции сохраняются все возможные варианты организации нервно-мышечных отношений и соответственно мотонейронных пулов спинного мозга, причем часто независимо от филогенетического уровня.
56