Для построения схем эволюционного развития ЦНС позвоночных необходим весь комплекс сведений об организации, нервной ткани: от анатомического описания отделов мозга до ультрамикроскопических, цитохимических и молекулярно-биологических данных. Все это дает возможность создать целостное представление об уровне организации и развития нервной: системы, оценить степень межвидовой и индивидуальной изменчивости структур нервных центров, уточнить "карты мозга".
Из множества нейрогистологических методов цитоархитектонический в наибольшей степени выявляет специфику мозга животных. Цитоархитектоника основана на анализе структурных различий в нервной ткани и использует для этого такие морфологические признаки, как величина и форма входящих в состав центра клеточных элементов, их число, плотность, наличие или отсутствие специализированных форм. Особое места отводится параметрам, связанным с пространственными, стереологическими характеристиками нервного центра: его объему, форме, характеру взаиморасположения клеток. Каждый участок мозга характеризуется определенным набором признаков. Значимость этих признаков различна: одни из них имеют принципиальное, первичное значение, рано появляются в онтогенезе и могут быть положены в основу разделения мозга на крупные области или зоны (например, при выделении формаций древней, старой и новой коры), другие, более частные, имеют вторичное значение, в онтогенезе появляются позднее и служат для выделения в составе основных подразделений более мелких структур (например, отделы коры, ее поля и подполя).
Кроме структурных признаков при морфологическом разделении мозга необходимо учитывать данные о его гистохимической природе, хемоархитектонике, системе афферентных и эфферентных связей (ходологии). Вместе с тем при проведении сравнительно-морфологического исследования и составлении цитоархитектонических карт мозга необходимо учитывать фактор внутри и межвидовой изменчивости, которая может достигать больших значений. Так, величины индивидуальной, внутривидовой, изменчивости в новой коре достигают 10 - 15%,
30
что вызывает определенные трудности при сопоставлении карт мозга.
В настоящее время, благодаря методам автоматизированного анализа, стало возможным получать количественную информацию о стуктуре мозга на качественно новом уровне. Особенно перспективен такой подход при изучении пространственных, стереологических, характеристик нервных центров.
Наиболее ярко эволюционные и адаптивные изменения нервных центров проявляются в структуре дендритных ветвлений нейронов. Поэтому вполне естественны попытки нейроморфологов создать классификацию нейронов, основанную на этом признаке. Так, Э. Рамон-Молинер предложил разделить все нейроны на три основных класса: изодендритные, аллодендритные и идиодендритные.
Класс изодендритных (неспециализированных) нейронов характеризуется относительно слабым развитием дендритнога поля: отростки не многочисленны, ветвятся слабо. В зависимости от их ориентации среди изодендритных нейронов выделены лептодендритные, лофодендритные и радиальные.
Класс аллодендритных (специализированных) нейронов отличается значительным усложнением системы дендритов. Это выражается в определенной ориентации отростков, появлении: специфических форм дендритного поля. К аллодендритным нейронам относятся, например, пирамидные нейроны коры и. многие нейроны таламических ядер.
Класс идиодендритных (высокоспециализированных) нейронов объединяет большую группу клеток различных отделок ЦНС, отличающихся высокодифференцированной системой дендритного ветвления. Морфологически это выражается в резком увеличении разветвленное™ отростков, в появлении узлов ветвления и системы возвратных коллатералей.
На основе данной классификации была составлена схема возможных преобразований нейронной структуры мозга позвоночных (рис. 11). Полагают, что для мозга гипотетического предка позвоночных характерны нейроны двух основных типов: лепто- и лофодендритные, расположенные в основном в перивентрикулярных областях мозга. В эволюции, вероятно, происходило изменение локализации этих двух семейств нейронов, что привело к появлению перивентрикулярных и субповерхностных слоев мозга. Максимального выражения этот процесс достигает у млекопитающих, где он проявляется в двух основных тенденциях усложнения нейронной структуры мозга - теленцефалической и стволовой. Теленцефалическая тенденция (описанная еще С. Рамон-и-Кахалем) проявляется в появлении большого количества специализированных типов нейронов
31
с определенно ориентированными дендритными стволами (например, гранулярные нейроны гиппокампа, пирамидные нейроны новой коры, клетки Пуркинье мозжечка, нейроны сетчатки и обонятельной луковицы). Вторая линия развития - стволовая - связана с преобразованием изодендритных нейронов ствола в систему высокоспециализированных, идиодендритных, клеток (например, "кисточковые" нейроны специфических центров ствола и промежуточного мозга). Следует отметить, что при этом значительная часть изодендригных клеток сохраняется в области осевой ретикулярной формации.
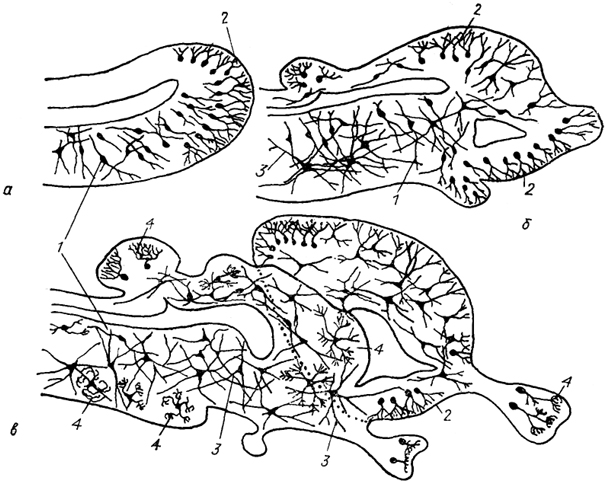
Рис. 11. Схема возможной эволюции нейронной структуры мозга позвоночных (Ramon-Moliner, 1975).
а - мозг гипотетического предка позвоночных; б, в - современных позвоночных низших (б) и высших (в). 1-4 - различные типы нейронов: лептодендритные (1), лофодендритные(2), аллодендритные (3), идиодендритные (4).
Классификация Рамон-Молинера в настоящее время широко используется нейроморфологами для изучения эволюционных преобразований структуры мозга. Более того, подобный подход позволяет провести теоретический анализ межнейронных взаимодействий в нервных центрах. Так, при равенстве линейных параметров дендритов (длина, диаметр) объем, занимаемый собственно дендритами, по отношению к объему дендритного поля нейрона будет наименьшим для изодендритных клеток, большим для аллодендритных и максимальным для идиодендритных клеток. Эта зависимость имеет важное значение
32
для понимания организации взаимоотношений нейронов с афферентными волокнами. Так, в нервном центре, образованном изодендритными клетками, будет больше контактов между
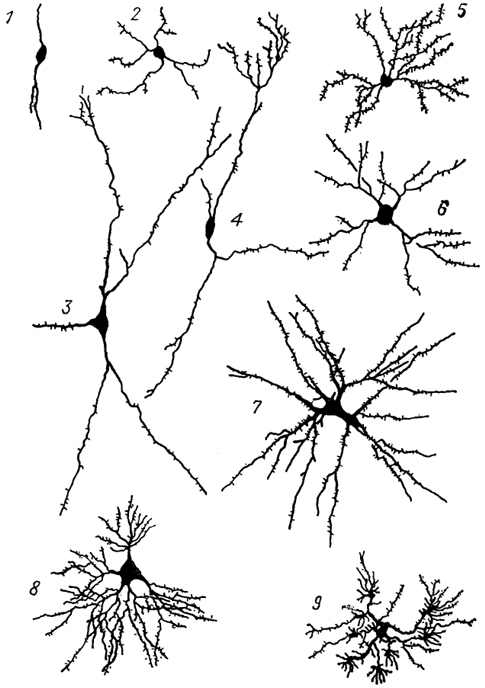
Рис. 12. Различные виды длинноаксонных нейронов подкорковых центров переднего мозга собаки (Леонтович, 1978).
1-3 - редковетвистые нейроны: нейробластоформный (1), короткодендритный (2), ретикулярный (3); 4-9 - густоветвистые нейроны: штамбовый (4), древовидные: густо-шипиковый (5) и редкошипиковый (6), мультиполярный гигантский (7), кустовидный (8) и кисточковый (9).
дендритами данной клетки и различными аксонами, входящими в этот центр, т.е. связи будут неспецифическими, случайными, тогда как в нейронных популяциях аллодендритного или идиодендритного типов будет больше контактов, образованных дендритами данной клетки с определенными аксонами, т.е. связи будут более специфическими.
Классификация Т. А. Леонтович акцентирует внимание на количественных характеристиках дендритной системы нейронов.
33
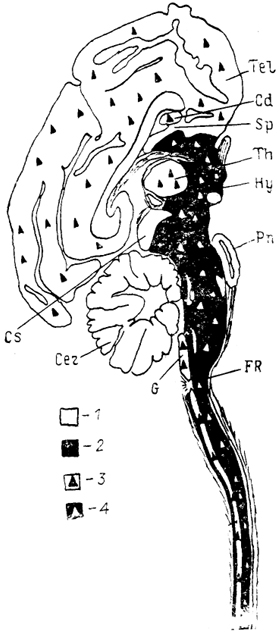
Рис. 13. Схема распределения элементов редковетвистой и густоветвистой нейронных систем в структурах головного мозга (Леонтович, 1978).
1, 2 - нейронные системы мозга: густоветвистая (1) и редковетвистая (2); 3 - ретикулярные клетки в структурах густоветвистой системы; 4 - густоветвистые (мультиполярные гигантские) клетки в структурах редковетвистой системы.
Все нейроны разделяют на две категории: длинноаксонные и короткоаксонные, в каждой из которых описано несколько классов клеток. Длинноаксонные нейроны разделяются на редко- и густоветвистые. Редковетвистые включают три класса клеток: нейробластоформные, короткодендритные и ретикулярные, а густоветвистые - пять классов: мультиполярные гигантские, штамбовые, древовидные, кустовидные и кисточковые (рис. 12). Каждый класс нейронов объединяет несколько разновидностей клеток и характеризуется определенным набором количественных признаков. Классификация Т.А. Леонтович разработана на мозге млекопитающих, однако она с успехом применяется для изучения нейронной структуры мозга позвоночных других групп, поскольку основана на универсальных, единых для всех нейронов, принципах ветвления дендритов и аксонов.
Исследование нейронной структуры мозга позвоночных различных рангов позволило выдвинуть и обосновать ряд общих принципов ее развития.
В мозге позвоночных не происходит "отрицания" старых форм и типов клеток, отделов. и даже целых систем мозга. В процессе эволюции они дополняются новыми образованиями, преобразуются или сохраняют принципы своего строения неизменными (принцип дополнительности). Это приводит к значительной гетерогенности нейронного состава многих отделов мозга, где наряду с эволюционно новыми, высокодифференцированными элементами часто сохраняются элементы, свойственные представителям более древних групп позвоночных. Об этом, в частности, свидетельствует факт "проникновения" ретикулярных редковетвистых нейронов - одной из наиболее филогенетически древних форм нейронов, характерной
34
для стволовых отделов мозга позвоночных - во все вышележащие отделы, включая конечный мозг (рис. 13).
Большое количество нейронов совмещает в себе черты строения разных клеточных типов, причем у высших позвоночных число таких "переходных" клеточных форм значительно возрастает (принцип структурной переходности). Значение их в функционировании нервных центров достаточно велико - благодаря сочетанию признаков, характерных для разных типов нейронов, они могут выполнять более широкие функциональные задачи, нежели крайние по форме нейронные типы.
На цитоархитектоническом уровне принцип структурной переходности проявляется в присутствии в ряде нервных центров (особенно в коре больших полушарий) межуточных формаций, соединяющих разные по строению и происхождению отделы. Данный принцип, в частности, свидетельствует о многоэтапности и неравномерности развития нервной системы в эволюции позвоночных.
Нейроны одного вида различаются между собой рядом признаков клеточной структуры, вследствие чего возможно определить лишь границы типовой формы нейронов (принцип полиморфизма). Так, даже у одного нейрона дендритные и аксонные ветви отличаются друг от друга по многим признакам, что обусловливает многообразие переходных форм нейронов. В процессе эволюции полиморфизм нейронной структуры мозга нарастает; причем это выражается, в частности, в усложнений ветвления отростков.
В нервной системе выделяются три основные группы синаптических контактов: электротонические, химические и смешанные, электро-химические. Они обнаружены практически во всех отделах ЦНС позвоночных разных групп и различаются между собой - в гомологичных центрах или нейронах - по форме синаптичеокого бутона, размеру, количеству и характеру распределения синапсов на теле и отростках нейронов.
Электротонические синапсы представляют собой особую разновидность щелевого межклеточного контакта, приспособленного для прямой передачи нервного электрического импульса с одной клетки на другую. Как и в других видах синапсов, здесь выделяют пре- и постоинаптическую части и синаптическую щель. Однако есть и специфические черты организаций: отсутствие субмембранных специализаций пре- и постсинаптических мембран, узкая, 2-3 нм, синаптическая щель, наличие в мембранах особых белковых комплексов - коннек-сонов, контактирующих в синаптической щели друг с другом и обеспечивающих проведение импульса, отсутствие, как правило, в пресинапсе синаптических пузырьков. Такая структура
35
контакта обусловливает его быстродействие, надежность в работе, практическую неутомляемость, возможность проведения импульса в обоих направлениях. В отношении последней характеристики необходимо подчеркнуть, что в ряде нейронных систем мозга позвоночных (например, в системе мюллеровских и маутнеровских клеток круглоротых и рыб) импульсы передаются в строго определенном направлении - от "командных" мюллеровских и маутнеровских нейронов. Электротонические синапсы обнаружены во многих отделах нервной системы у всех изученных представителей позвоночных.
Таблица 1. Механизмы передачи в гомологичных синапсах ЦНС (Шаповалов, Ширяев, 1987)
| Класс позвоночных |
Нейроны, образующие синапс |
| первичный афферент-мотонейрон |
ретикулоспинальный нейрон-мотонейрон |
вестибулярный афферент-ядро Дейтерса |
Круглоротые:
минога |
Смешанный |
Смешанный |
Смешанный |
Амфибии:
лягушка |
-"- |
Химический или смешанный |
Химический или смешанный |
Млекопитающие:
крыса
кошка |
Химический
-"- |
Химический
-"- |
-"-
Химический |
Химические синапсы характеризуются наличием в пресинапсе синаптических пузырьков, развитой системой пре- и постсинаптической субмембранных специализаций ("а основании этого выделяют две разновидности синапсов - симметричные и асимметричные), достаточно широкой синаптической щелью (от 20 до 50 нм). Показано, что любая часть нейрона - тело, аксон, дендриты - может образовывать синапсы. Наибольший интерес представляют аксо-аксонные и дендро-дендритные контакты, поскольку они имеют важное значение для обеспечения процессов авторегуляции деятельности нейронов (при взаимодействии аксонных коллатералей с аксонным холмиком собственного нейрона) к для осуществления тормозных взаимодействий в нейронных цепях. Очень часто соединения подобных типов являются контактами реципрокными, т. е. имеющими по две пре- и постсинаптические зоны, что обеспечивает двустороннее проведение импульса. Встречаются и так называемые смешанные, электро-химические, контакты.
Вопрос о происхождении различных типов синапсов в гомологичных нервных центрах мозга позвоночных разного филогенетического уровня не решен. Одни исследователи рассматривают электротонические синапсы как первичные в эволюции нервной системы и указывают на существование в филогенезе процесса замещения электротонических контактов химическими
36

Рис. 14. Синаптические комплексы различного вида в структурах нервной системы млекопитающих.
а-е - дивергентные комплексы в структурах моста (а), кохлеарном ядре (б), вентробазальном таламусе (в), коре мозжечка (г), желатинозной субстанции спинного мозга (д), клиновидном ядре продолговатого мозга (е); ж, з - конвергентные комплексы на нейронах таламуса (ж) и мотонейронах спинного мозга (з). 1 - аксоны, 2 - дендриты, 3 - дендритные шипики. Стрелки - тормозные воздействия.
(табл. 1). Другие считают, что как электротонические, так и химические синапсы возникли в эволюции нервной системы очень рано и имеют широкое распространение в нервных центрах мозга позвоночных всех филогенетических групп. Преобладание в ряде нервных центров того или иного типа синаптической
37
передачи определяется функциональными задачами данного центра, а не его происхождением. Эта точка зрения базируется на фактах обнаружения типичных химических и электротонических синапсов в нервной системе низших многоклеточных животных с одной стороны, и в высших интегративных центрах конечного мозга млекопитающих и птиц, с другой. Более того, показано, ч го синапсы, образуемые одним аксоном на разных постсинаптических элементах, могут работать с использованием и химического, и электротонического механизмов.
Связи между нейронами могут осуществляться как одиночными синапсами различного вида, так и сложными комплексами типа "гломерул" (рис. 14). Синаптические комплексы построены по двум основным принципам: конвергенции и дивергенции. В центре конвергентного синаптического комплекса находится один постсинаптический элемент - тело или отросток нейрона - на котором оканчивается несколько пресинапсов различного происхождения и типа. В дивергентном комплексе отношения обратные: одна пресинаптическая часть контакта - чаще всего окончание или расширение аксона - оказывает влияние на несколько постсинаптических элементов.
В последние годы во многих отделах нервной системы позвоночных обнаружены так называемые модуляторные контакты. Нейроактивная субстанция в них заключена, как правило, в крупные секреторные гранулы. Путем экзоцитоза она выделяется в межклеточные пространства и, достигая рецепторов постсинаптических клеток, воздействует не на ионную проницаемость мембраны (как в типичном химическом синапсе), а на синтетические процессы в клетках-мишенях, изменяя, в частности, уровень синтеза цАМФ, активность мембранной АТФазы или протеинкиназ. Имеется еще ряд существенных отличий в организации и функционировании синаптических и модуляторных контактов в нервной системе позвоночных (табл. 2).
Важно отметить, что термины "нейромедиатор", "нейромо-дулятор", "нейрогормон" отражают скорее механизм взаимодействия этих соединений с клетками-мишенями, чем их химическую природу. Одно и то же вещество в разных отделах нервной системы организма может выступать как в роли медиатора, так и нейромодулятора или нейрогормона. Многие нейропептиды, например энкефалины и эндорфины, ранее рассматриваемые только как нейрогормоны, выделяются аксонными терминалями и выступают в роли типичных нейромедиато-ров. И наоборот, отдельные биогенные амины (адреналин, дофамин, серотонин) не только действуют через межнейронные синапсы, но могут также выделяться нейросекреторньши клетками гипоталамических ядер или органов циркумвентрикулярной системы в кровь и цереброспинальную жидкость, распространяясь
38
затем в организме на значительные расстояния и действуя как типичные нейрогормоны.
Таблица 2. Различия между контактами в ДНС позвоночных (по: Vizi, 1984)
| Параметры |
Контакты |
| синаптические |
модуляторные |
нейросекреторные* |
| Тип связи |
Один на один |
Один на много |
Один на много |
| Время действия |
Короткое, мс |
Длительное, с |
Длительное, мин и ч |
| Механизм действия |
На ионную проницаемость мембран |
Без прямого действия на ионную проницаемость мембран |
|
| Характер действия |
Фазный |
Тонический |
Тонический |
| Место действия |
Пре- и пост-синапс |
Пре- и пост-синапс |
Пре- и пост-синапс, клетки-мишени, органы |
| Место выведения |
Синаптическая щель (5 - 50 нм) |
Межклеточные пространства, мкм, мм |
Межклеточные пространства, кровь, спинномозговая жидкость |
| Активность соединений |
Низкая |
Средняя |
Чрезвычайно высокая |
| Специфичность соединений |
Высокая |
Высокая |
Высокая |
| Скорость синтеза |
Средняя |
Средняя |
Низкая |
| Молекулярный вес |
Небольшой |
Небольшой |
Небольшой и средний |
| Концентрация молекул |
Высокая(10-4 - 10 3M) |
Средняя(10 6 - 10-5M) |
Низкая(10-9 - 10-12M) |
*Нейросекреторные окончания образованы отростками специализированных клеток, составляющих важный интегрирующий компонент нервной системы. Нейросекреторные клетки - филогенетически древние элементы нервной системы, сочетающие в себе свойства нервных и секреторных клеток. Они синтезируют и выделяют целый комплекс биологически активных веществ - нейрогормонов. В настоящее время описано несколько десятков соединений, в основном пептидной природы, выполняющих эту важную функцию.
С помощью биохимических, авторадиографических и иммуноцитохимических методов в ЦНС выявлено несколько десятков таких соединений. При этом нейроны могут синтезировать и выделять в своих терминалях целый комплекс посредников (до 12!). В исследованиях на культуре нейронов показано, что клетки синтезируют несколько физиологически активных межнейронных посредников, сочетание которых в разных нейронах варьирует. Так, в 217 изученных нейронах было отмечено 20 комбинаций из 4 идентифицированных веществ, причем при одинаковом наборе посредников их количественное соотношение оказалось разным.
Способ накопления веществ в одном синапсе может быть различным: в одних и тех же синаптических пузырьках, в разных
39
популяциях пузырьков и секреторных гранул, в цитоплазме аксонной терминалы и пузырьках. В связи с этим наиболее сложным в проблеме межнейронных взаимодействий является исследование механизма действия различных субстанций, выделяемых одной аксонной терминалью. Существует два основных варианта действия медиаторов: дивергентный и независимый, при котором каждый выделяемый из терминали медиатор действует на определенную популяцию клеток-мишеней; и конвергентный и зависимый, при котором один медиатор оказывает определенное влияние на взаимодействие постсинапти-ческой клетки с другим медиатором на разных этапах синаптической передачи.
Таким образом, краткий обзор строения и функций синаптических контактов в ЦНС позвоночных показал, что на фоне относительного единообразия структурной организации синапсов и консервативности химических соединений, используемых для передачи нервного импульса, в нервных центрах имеется возможность их эволюционного развития и усложнения путем комбинации различных типов межнейронных соединений (принцип полиморфизма) и использования различных посредников - медиаторов, модуляторов, нейрогормонов - в организации нейронных цепей.
Идея о том, что нервные центры состоят не из отдельных цепочек нейронов, работающих независимо друг от друга, а организованы в блоки или модули, объединяющие от нескольких клеток до нескольких тысяч нейронов, которые функционируют как единое целое, была выдвинута С. Рамон-и-Кахалем еще в начале XX в.
В ЦНС позвоночных модули были впервые обнаружены и описаны в коре мозжечка с его упорядоченным расположением •слоев и клеточных элементов. Позднее на тангенциальных срезах соматосенсорной коры мозга мыши были выявлены кольцеобразные скопления нейронов, названные "бочонками" (англ, barrels). Оказалось, что число и пространственное расположение этих модулей точно соответствует числу и расположению-чувствительных вибрисс на морде животного. Это явилось одним из первых доказательств того, что модуль является структурно-функциональной единицей нервного центра. В дальнейшем модули были обнаружены в разных отделах ЦНС у многих позвоночных. Пространственная организация модулей может быть различной: от плоских дисков, характерных для нейропиля спинного мозга, до сложнейших пространственных комплексов- "бочонков" в новой коре и стриосом в базальных ядрах конечного мозга. Однако независимо от конкретного вида модуля, он представляет собой популяцию нейронов, способных
40
к возбуждению или торможению относительно независимо от подобных процессов в соседних группах нейронов. В состав любого модуля входят три группы клеток: нейроны, принимающие афферентные воздействия, эфферентные клетки, а также нейроны, осуществляющие локальные связи. Межнейронные контакты внутри модуля осуществляются системой "микромодулей", структурной основой которых могут быть пучки дендритов и аксонов.
Размеры и количество модулей в разных отделах ЦНС и у разных животных колеблются в широких пределах. Так, в новой коре млекопитающих насчитывается от 0,6 до 2-3 млн. объединений нейронов, в составе которых находится от 3 до 10 тыс. клеток. Размеры модуля определяются объемом ветвления афферентов, входящих в данный модуль. Элементарные модули могут объединяться в более сложные комплексы, содержащие до нескольких десятков модулей. Морфологической основой такого рода объединения могут служить аксонные коллатерали нейронов модуля, распространяющиеся на большие расстояния (в новой коре от 3 до 8 мм).
Таким образом, центральная нервная система всех позвоночных имеет единый план строения, основанный на общих принципах формирования и развития нервных центров всех уровней организации.
41
