Представленные ниже описание и схема развития системы мозговых желудочков (рис. 8) получены при исследовании онтогенеза ЦНС у представителей млекопитающих. На стадии 20 сомитов (у человека приблизительно 24-й день развития) в головном отделе нервной трубки формируются три первичные мозговые полости (стадия трех мозговых пузырей), полость первичного переднего мозга (prosocoele), средняя мозговая полость (mesocoele) и полость мозгового ствола (rhombocoe1е). Остальная часть полости нервной трубки на этой стадии получила название syringocoele; на ее основе в дальнейшем образуется центральный спинномозговой канал.
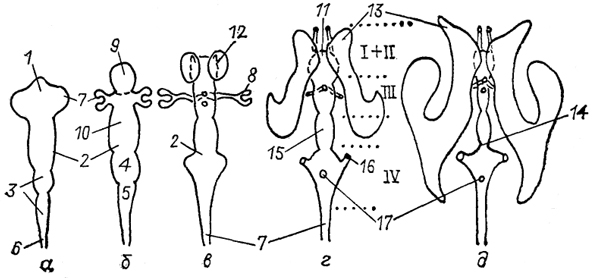
Рис. 8. Схема развития мозговых желудочков млекопитающих (McFarland е. а., 1969).
а-д - этапы развития мозговых желудочков на стадии трех (а) и пяти (б, в) мозговых пузырей, в пре- (г) и постнатальный (д)периоды. 1 - прозоцель и его производные: телоцель (9) и диоцель (10); 2 - мезоцель; 3 - ромбоцель и его производные: метацель (4) и миелоцель (5); 6 - сирингоцель и его производное - спинномозговой канал (7); 8 - глазной пузырь; 11, 12 - полость (11) и отверстия (12) Монро; 13 - боковые желудочки (I, II); 14 - перешеек; 15 - водопровод мозга; 16, 17 - мозговые отверстия Люшка (16) и Мажанди (17). Пунктиром обозначены границы I-IV мозговых желудочков.
Первичный конечный мозг развивается как вырост первого мозгового пузыря ростральнее оптической зоны (стадия пяти мозговых пузырей). Формирующаяся при этом борозда отделяет ростральную часть переднего мозга с его полостью (telocoele medialis) от каудальной части переднего мозга с его полостью (diocoele), из которой в дальнейшем образуется промежуточный мозг. Полость среднего мозгового пузыря на этой стадии еще превосходит по размерам полость переднего мозга. Полость ствола мозга разделяется на полости заднего (metacoele) и продолговатого (myelocoele) мозга.
На стадии 31 сомита в полости конечного мозга (telocoele) формируются билатеральные выпячивания, образующие боковые
23
желудочки, ventriculi laterales, которые отделяются от непарной части полости конечного мозга септальной бороздой. Полость непарной части конечного мозга в дальнейшем становится полостью Монро, которая соединяется с III мозговым желудочком (полостью промежуточного мозга) межжелудочковыми отверстиями, или отверстиями Монро. В то же время в развивающихся полушариях образуется еще одна борозда - ринальная, которая отделяет зачатки будущих обонятельных луковиц от остальной части полушарий.
В результате быстрого роста дорсального отдела таламуса латеральные стенки полости промежуточного мозга сближаются, образуя узкую щель - III мозговой желудочек, ventriculus tertius. Следует сказать, что данный процесс наблюдается лишь у высших млекопитающих, тогда как у всех остальных позвоночных объем III желудочка остается достаточно большим. Полость среднего мозга на этой стадии развития также еще большая и только впоследствии сужается, формируя водопровод мозга, aqueductus cerebri. В заднем мозговом пузыре образуются латеральные выросты, на концах которых формируются отверстия Люшка и Мажанди, играющие важную роль в циркуляции цереброспинальной жидкости. Отверстие Люшка, в отличие от отверстия Мажанди, есть у всех млекопитающих. У человека отверстие Мажанди на 5 - 6-м месяце эмбрионального развития открывается в полость мозговых оболочек.
Понимание процесса развития и изучение конфигурации мозговых желудочков в филогенезе позвоночных позволяют лучше ориентироваться при описании структур мозга (рис. 9). При этом важное значение приобретает проблема гомологии отдельных образований желудочковой системы у представителей разных групп. Даже в пределах одного класса, например млекопитающих, имеется значительная вариабельность в развитии тех или иных отделов.
Все мозговые желудочки и спинномозговой канал выстланы изнутри слоем специализированных эпендимных клеток, являющихся разновидностью глиальных элементов нервной ткани и происходящих из матричных клеток нервной трубки. Однако эпендимная выстилка мозговых желудочков неоднородна, в ней есть участки, имеющие атипическое строение - так называемые органы циркумвентрикулярной системы. Наиболее подробно она изучена у млекопитающих, где выделено несколько групп таких органов (рис. 10).
К первой группе органов циркумвентрикулярной системы принадлежат участки специализированной эпендимной выстилки, образующие сосудистые сплетения (PC III, PC IV), а также структуры парафиза (РА), дорсального и сосудистого мешков III мозгового желудочка.
Сосудистые сплетения находятся в полостях боковых и IV мозгового желудочков. В них происходит образование основного
24
объема цереброспинальной жидкости. Эпендимные клетки сплетений часто называют хориоидным эпителием. Они имеют кубовидную форму и на апикальной поверхности несут многочисленные микроворсинки. Плотные контакты между эпендимными клетками препятствуют прямому проникновению веществ из кровяного русла в полость желудочков.
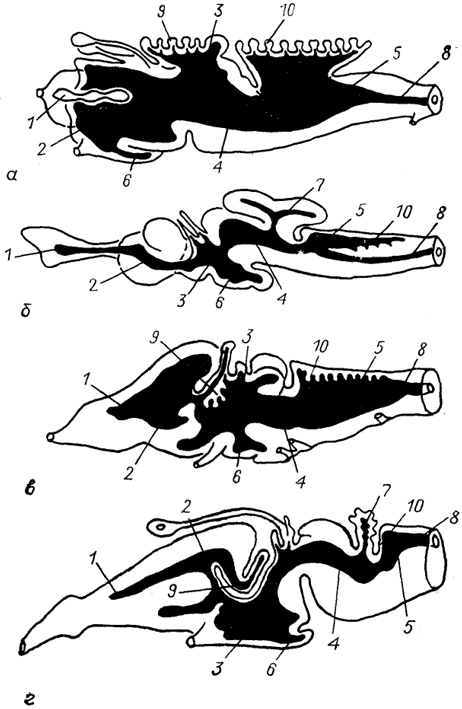
Рис. 9. Строение мозговых желудочков у ряда позвоночных (McFarland e. а., 1969).
а - минога, б - акула, в - лягушка, г - ящерица. 1 - обонятельные желудочки; 2-5 - латеральные (2), третий (3), среднемозговой (4), четвертый (5) желудочки; 6-8 - полость воронки гипоталамуса (6), мозжечка(7), спинного мозга(8); 9, 10 - сосудистые сплетения в III (9) и IV (10) мозговых желудочках.
Цереброспинальная жидкость (ЦСЖ) играет роль гидростатического
25
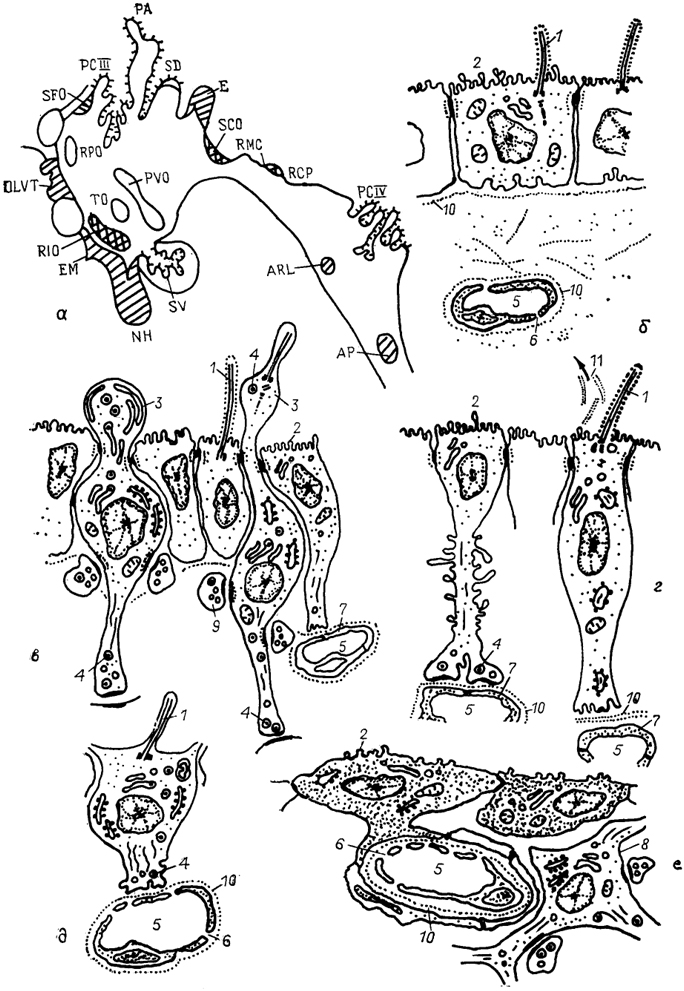
Рис. 10. Циркумвентрикулярная система мозга позвоночных (по: Oksche, 1973, 1988).
а - общий план строения; б-е - фрагменты ультраструктуры клеток эпендимы сосудистого сплетения (б), ликворконтактирующих нейронов преоптической зоны (в), танницитов субкомиссуального органа (г), пинеалоцитов эпифиза (д) и специализированной эпендимы area postrema (е). 1 - реснички; 2 - микроворсинки; 3 - выросты апикальной части клеток; 4 - секреторные гранулы и пузырьки; 5 - кровеносные капилляры; 6, 7 - эндотелий сосудов с порами (6) и без них(7); 8 - нейрон; 9 - синапсы; 10 - базальная мембрана; 11 - рейснеровские волокна. Объяснения в тексте.
26
буфера, заполняя пространства между мозговыми оболочками; поддерживает ионный баланс в мозговой ткани; является средой-переносчиком биологически активных веществ: нейромедиаторов, гормонов, нейросекретов; удаляет из нервной ткани продукты метаболизма, попаданию которых в кровь препятствует гемато-энцефалический барьер. Продуцирование ЦСЖ является активным процессом. Ее отток из желудочков осуществляется через полость ромбовидного мозга в подпаутинное пространство мозга через отверстия Люшка и Мажанди, а оттуда - в венозные синусы твердой мозговой оболочки.
Среди низших позвоночных степень развития сосудистых сплетений сильно варьирует. У круглоротых в переднем мозге они практически не развиты, что компенсируется значительным развитием сосудистого сплетения продолговатого мозга (рис. 9). У рыб выстилка сосудистых сплетений представлена эпендимными клетками самых разных типов: от кубовидных клеток, связанных с богатой сетью капилляров мягкой мозговой оболочки, до уплощенных клеток без ресничек и микроворсинок, фактически не участвующих в образовании ЦСЖ (например, в конечном мозге некоторых бесхвостых амфибий и костистых рыб).
Парафиз (РА) - наименее изученный орган циркумвентрикулярной системы. У некоторых хрящевых рыб, например акул и химер, парафиз имеет дольчатое строение, а цилиндрические эпендимные клетки, в отличие от аналогичных элементов сосудистых сплетений, лишены ресничек. У высших представителей хрящевых рыб (в том числе и высших акул) парафиз, по-видимому, дегенерирует. У костных рыб степень развития парафиза варьирует от весьма значительной (осетровые и ильные рыбы) до крайне слабой (представители некоторых семейств высших костистых рыб). Парафиз у амфибий выражен хорошо, у высших позвоночных - умеренно, а у млекопитающих во взрослом состоянии часто отсутствует.
Таким образом, парафиз хорошо развит у примитивных групп, как у рыбоподобных, так и у тетрапод, у высших же представителей этих групп он менее выражен. Происхождение парафиза неясно: одни авторы считают его самостоятельным отделом циркумвентрикулярнок системы, другие - производным сосудистых сплетений.
Сосудистый мешок (SV)найден практически у всех хрящевых и костных рыб, за исключением отдельных представителей,
27
где его отсутствие, по-видимому, вторично. У костистых рыб степень его развития обусловлена их образом жизни и средой обитания. Так, у пресноводных видов сосудистый мешок слабо-дифференцирован, тогда как у эвригалинных (например, Апguella, Hypomesus) он выражен очень хорошо. Однако такая корреляция не прослеживается у представителей других групп рыб: у ряда пресноводных хрящевых рыб, костных ганоидов и брахиоптеригий эта структура развита весьма значительно. Другая тенденция в эволюции сосудистого мешка заключается в его постепенной изоляции от III желудочка. Это направление развития четко прослеживается от низших рыб к высшим, а у некоторых костистых рыб (например, представителей отряда Sardinella) он вообще замкнут. Еще одна особенность строения сосудистого мешка связана с распределением участков специализированной эпендимы: у хрящевых и костных ганоидов она обычно находится только в зоне, прилежащей к нейроги-пофизу, а у представителей высших отрядов костистых рыб образует всю выстилку сосудистого мешка. Неясно, имелся ли этот орган у предков позвоночных, а следовательно, остается открытым вопрос: первично или вторично его отсутствие у современных круглоротых?
Во второй группе органов циркумвентрикулярной системы выделяют две подгруппы. В первую входят субкомиссуральный орган (SCO), выросты эпендимной выстилки желудочка среднего мозга (RMC, RCP) и воронка гипоталамуса (RIO). В этих органах выстилка полости мозговых желудочков представлена участками специализированной эпендимы, клетки которой обладают ярко выраженной секреторной активностью. Эти клетки, получившие название "эпендимные таннициты", имеют как апикальные отростки, контактирующие с полостью желудочка", так и базальные отростки, заполненные секреторными гранулами и достигающие капилляров (рис. 10). Ко второй подгруппе относятся паравентрикулярный орган (PVO) и преоптический вырост (RPO), где наряду с танницитами в просвет желудочков выходят отростки специализированных нейросекре-торных клеток преоптического и паравентрикулярного ядер гипоталамуса. Ликворконтактирующие нейроны этих образований секретируют в цереброспинальную жидкость биологически активные вещества - нейрогормоны, медиаторы, нейропептиды, которые затем через систему мозговых оболочек попадают в кровь.
Субкомиссуральный орган (SCO)обнаружен практически у всех представителей позвоночных и является, по-видимому, одним из самых филогенетически древних отделов циркумвен-трикулярной системы. У круглоротых SCO парный, в то время-как у других позвоночных парными являются только его ростральные части. В некоторых группах млекопитающих (насекомоядные, китообразные, высшие приматы) с возрастом наблюдается
28
некоторая редукция SCO. Клетки этого органа отличаются ярко выраженной полярностью. Секрет выводится в ЦСЖ в виде фибриллярного материала - так называемых Рейснеровских волокон. Эти волокнистые структуры накапливаются в каудальной части центрального спинномозгового канала или в субарахноидальном пространстве в области задней комиссуры, откуда активно всасываются в кровь. Предполагают, что секрет SCO представляет собой гормон, участвующий в регуляции процессов роста и дифференцировки организма. SCO в онтогенезе позвоночных начинает функционировать очень рано и наиболее активен в первую половину эмбрионального развития.
Третья группа органов цирку мвентрикулярной системы представлена такими участками стенок мозговых желудочков, где эпендимная выстилка полностью или частично замещается на нервные или нейросекреторные клетки и имеется необычная система капилляров. К такого рода участкам относятся: субфорникальный орган (SFO), сосудистый орган конечной пластинки (OVLT), срединное возвышение (ЕМ), нейрогипофиз (NH), эпифиз (Е), расположенные в III желудочке, и зона латеральных выростов и area postrema (ARL, АР), расположенные в полости IV мозгового желудочка (рис. 10).
В субфорникальном органе (SFO) капилляры непосредственно контактируют с эпендимной выстилкой. У круглоротых и рыб SFO практически отсутствует, хотя участки, где крупные капилляры подходят к эпендиме, встречаются у многих костистых рыб в разных отделах мозга. Ряд исследователей связывает появление SFO в эволюции позвоночных с наземным образом жизни. Он хорошо развит у хвостатых амфибий, довольно слабо - у рептилий и птиц. У млекопитающих SFO сложно дифференцирован и включает в свой состав несколько типов нейронов, контактирующих с ЦСЖ и периваскулярным пространством капилляров. Полагают, что SFO принимает участие в регуляции водно-солевого обмена.
Area postrema (АР) - крупный орган циркумвентрикулярной системы, лежащий на дорсальной поверхности стенки желудочка, в каудальной части продолговатого мозга. АР представляет собой участок специализированного гематоэнцефалического барьера. Ее элементы выполняют нейросекретор-ную функцию, выделяя физиологически активные субстанции типа серотонина, энкефалина, холецистокинина и др., контролируют обмен натрия, регулируют кровообращение, дыхание, осморегуляцию, выделение и энергетический обмен. Эпендимные клетки АР лишены ресничек и связаны друг с другом плотными контактами. В паренхиме органа присутствуют группы глиальных клеток, нейронов и аксонных терминалей разного типа. Капилляры имеют многочисленные поры и окружены широким периваскулярным пространством, которое ограничено
29
плотной оболочкой из тел и отростков глиальных клеток (рис. 10).
Area postrema традиционно рассматривалась как область, присутствующая только в мозге млекопитающих и птиц. Однако современные исследования показали, что у многих представителей низших позвоночных она тоже имеется, и ее наличие или отсутствие не связано ни с филогенетическим положением, ни с- образом жизни животного.
30
