В трактовке морфофункциональной организации конечного мозга амниот, особенно рептилий и птиц, произошли значительные изменения. Ряд крупных образований, занимающих в полушарии субвентрикулярное положение и рассматриваемых ранее как субпаллиальные, на основании современных данных по эмбриогенезу, гистохимии, нейронной структуре и связям относят к отделам паллиума.
В конечном мозге рептилий паллиальными формациями являются собственно паллиальная (корковая) пластинка, разделенная на медиальный (гиппокампальный), дорсальный (общий) и латеральный (пириформиый) отделы, а также структуры так называемого дорсального вентрикулярного края (рис. 87).
Паллиум у разных рептилий имеет различную степень дифференцировки. У ящериц границы его отделов определяются достаточно четко по наличию зон перекрытия между медиальным и дорсальным и между дорсальным и латеральным паллиумом. У черепах, крокодилов и змей такого рода переходных зон нет и границы отделов паллиума определяются по изменению их внутренней структуры (цитоархитектонике, нейронному составу и связям).
В медиальном паллиуме большинства рептилий выделяют два участка: вентральный, мелкоклеточный (Ml), и дорсомедиальный, крупноклеточный (М2), составленные несколькими типами нейронов, в том числе пирамидоподобными. Источниками афферентов являются структуры конечного мозга (дорсальный и латеральный паллиум, ДВК, септум), переднее дорсолатеральное ядро таламуса, мамиллярные тела, ядра шва. Эфферентные проекции направляются в септум, амигдалу, передние отделы таламуса, гипоталамус. Через переднюю комиссуру медиальный паллиум связан с одноименной зоной паллиума другого полушария, а по системам внутриполушарных связей - с дорсальным паллиумом и ДВК.
Дорсальный паллиум рептилий разделяется на дорсомедиальную и дорсолатеральную части. Дорсомедиальная сходна у всех рептилий по структуре и связям, дорсолатеральная различна у чешуйчатых рептилий и черепах. У последних ее нейроны получают проекции по системе латерального пучка переднего мозга от дорсальной части ЛКТ и из вентрального таламуса, тогда как у ящериц в дорсолатеральный отдел дорсального паллиума
255
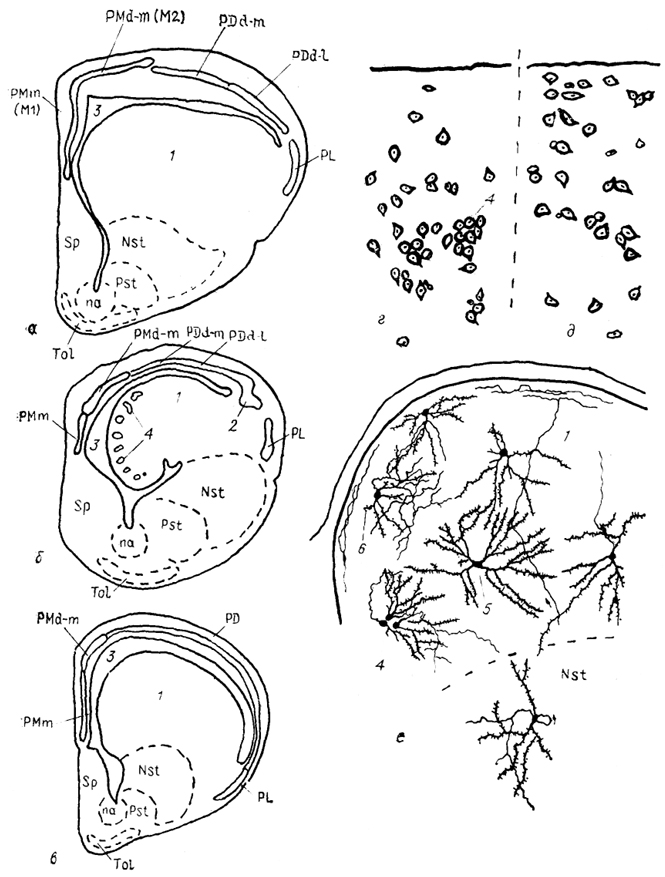
Тис. 87. Структура конечного мозга чешуйчатых рептилий
(а), черепах
(б) и крокодилов
(в), цитоархитектоника дорсо-вентрикулярного края (ДВК) первого
(г) и второго
(д) типов и фрагмент нейронной структуры ДВК гадюки
(е) (Обухов, Санькова, 1986; Ulinski, 1990).
1 - ДВК, 2 - паллиальное утолщение, 3 - латеральный мозговой желудочек, 4 - клеточные группы, 5 - радиальный густоветвистый и 6 - короткоаксонный нейроны ДВК.
256
по системе медиального пучка проецируется только дорсолатеральное ядро таламуса. Другие афференты дорсального паллиума приходят из ростральных отделов ДВК, паллиально-го утолщения, гипоталамуса, вентральной тегментальной области, ядер шва и голубого пятна.
Основная часть эфферентов направлена в гипоталамус, медиальный паллиум, септум. Отмечены проекции в стриатум, ДВК, амигдалу, переднее обонятельное ядро, основную и добавочную обонятельные луковицы, дорсальный таламус (дорсомедиальное и дорсолатеральное ядро). Комиссуральные связи обеспечивает передняя комиссура.
Самый латеральный отдел дорсального паллиума образует так называемое паллиальное утолщение специфического строения. Корковая пластинка утрачивает здесь слоистую структуру, а нейроны группируются в несколько крупных скоплений. Наиболее выражено паллиальное утолщение у черепах. В эту область проецируются волокна из дорсального таламуса. Так, через круглое ядро в паллиальное утолщение мозга ящериц поступает зрительная информация. Высказано предположение, что у рептилий в эволюции сформировалось несколько самостоятельных таламо-кортикальных путей в медиальный и дорсальный паллиум, паллиальное утолщение, ДВК.
Долгое время дорсальный паллиум (особенно его латеральные участки) рассматривали как возможный гомолог специфических сенсорных зон неокортекса млекопитающих. Однако система двусторонних связей с медиальным паллиумом и септумом, а также наличие обонятельных проекций из латерального паллиума, свидетельствуют о том, что этот отдел полушария (по крайней мере, у ящериц) наряду с медиальным паллиумом является теленцефалическим компонентом лимбической системы.
Латеральный паллиум рептилий занимает часть латеральной стенки полушария и представлен популяцией радиальных и горизонтальных нейронов изодендритного типа. Он имеет, как и другие отделы паллиума, отчетливое корковое строение. Латеральный паллиум получает прямые обонятельные проекции. Остальные афференты приходят из ростральных отделов ДВК, амигдалы и ядра латерального обонятельного тракта. Эфференты направляются в дорсальный паллиум и мелкоклеточную часть медиального паллиума, где оканчиваются на базальных дендритах ее нейронов, а также в амигдалу.
Дорсальный вентрикулярный край (ДВК) есть в полушариях конечного мозга всех рептилий и состоит из двух отделов: переднего и заднего. Передний ДВК относится к собственно паллиальным формациям, а задний, включающий такое крупное образование, как сферическое ядро, - к амигдалярному комплексу.
Размеры и структура переднего ДВК у рептилий, как из разных групп, так и у близкородственных видов, сильно варьирует.
257
Так, у ящериц Dracomorpha ДВК Занимает от 22 до 44%, объема всего полушария, а у ящериц Lacertomorpha - о,т 7 до" 12%. Цитоархитектонически выделяют два типа ДВК- К первому типу относят ДВК, в составе которого выделяют поверхностную клеточную пластинку, образованную скоплением групп нейронов, и центральное ядро, 'имеющее диффузное строение (рис. 87). Такой тип ДВК отмечен у черепах, гаттерий, ряда змей и некоторых других чешуйчатых (сем. Gekkonidae, Lacertidae, Anguidae). В структуре ДВК второго типа отсутствует четкое разделение на клеточную пластинку и центральное ядро. Нейроны, как одиночные, так и в группах, равномерно распределены по всему ДВК. Этот тип ДВК встречается у крокодилов, некоторых змей и ящерицевых (сем. Agamidae, Iguanidae, Varanidae).
В ДВК рептилий обнаружены дискретные зоны проекций таламических ядер. Так, у безногой ящерицы желтопузика через, круглое ядро таламуса в латеральную зону ДВК поступает зрительная информация, а через nucl. reuniens в медиовентральную зону ДВК - соматическая и слуховая. На границе этих зон выявляется большое число полимодальных нейронов, что связано с явлением взаимопроникновения дендритов нейронов этих зон, где на них оканчиваются афференты другой модальности.
Подобные проекционные зоны обнаружены в мозге черепах и крокодилов, но, в отличие от ящериц, у них происходит обособление центральной зоны ДВК, получающей через медиальное заднее ядро таламуса соматическую информацию. Помимо проекций из таламуса, ДВК имеет обширные афферентные и эфферентные связи с другими отделами паллиума, стриатумом, амигдалой.
Таким образом, ДВК конечного мозга рептилий - высокодифференцированный отдел полушария, имеющий паллиальное происхождение. По наличию модальноспецифических сенсорных зон он сходен с неокортексом мозга млекопитающих, однако в отличие от ряда полей неокортекса, ДВК рептилий не имеет прямых проекций в моторные центры ствола и спинного мозга. Исследователи считают ДВК структурой, специфичной для конечного мозга рептилий, эволюция которой проходила независимо и параллельно развитию других паллиальных формаций мозга высших позвоночных.
К субпаллиуму конечного мозга рептилий относят стриатум и ряд структур вентромедиальной стенки полушария: обонятельный бугорок, септум, nucl. accumbens, и амигдалу, ядра которой расположены в вентролатеральной зоне полушария.
Стриатум разделяется на два отдела: дорсальный, получивший название неостриатум, neostriatum, и вентральный - палеостриатум, paleostriatum (рис. 87). Неостриатум составляет незначительную часть стриатарного комплекса и хорошо выражен у всех рептилий. Размеры и топография неостриатума подвержены довольно слабым межвидовым колебаниям. В его составе:
258
не выделяется каких-либо отдельных образований, а по уровню нейронной дифференцировки он значительно уступает соседнему ДВК. Палеостриатум всех изученных рептилий разделяется на две части: медиальную (мелкоклеточную) и латеральную (крупноклеточную).
Среди проекций неостриатума наиболее важными являются двусторонние связи с дорсальным паллиумом и передним ДВК, аналогичные неостриато-корково-неостриатарным связям конечного мозга млекопитающих. Обнаружены также двусторонние связи стриатума с тегментумом среднего мозга (ядром ТРС)Г которые сравниваются со стриато-нигро-стриатарными связями млекопитающих. Кроме того, показано, что проекции сенсорных, таламических ядер, проходящие в паллиум и ДВК, дают в стриатуме коллатерали (рис. 88). Основная масса эфферентных волокон неостриатума проходит в палеостриатум, который в свою очередь образует главные эфферентные тракты стриатума. Так, у ящерицы Gekko gekko были отмечены проекции в передние и задние энтопедункулярные ядра, в вентромедиальное и вентро-латеральное ядра таламуса. У черепах и крокодилов (кайманы) палеостриатум связан с ядром задней комиссуры, которое проецируется в тектум. Эфферентные проекции палеостриатума рептилий прослеживаются также в такие структуры, как ЦСВ среднего мозга, ядра шва, интерколликулярное ядро, латеральное ядро мозжечка и ретикулярную формацию.
Таким образом, структура и система связей стриатума позволяют рассматривать его как сложный корреляционный центр полушарий рептилий. Особо следует отметить формирование впервые у позвоночных системы двусторонних стриато-паллиальных связей.
Септум мозга разных рептилий имеет единый план строения, сходную нейронную организацию и состоит из медиального и латерального ядер. В септуме описано два основных типа нейронов: мультиполярные редковетвистые и веретеновидные биполярные. Главными проекциями септума являются билатеральные двусторонние связи с медиальным паллиумом и контрлатеральные афференты из дорсального паллиума, обонятельного бугорка и амигдалы. По медиальному обонятельному тракту ипсилатерально в септум приходят волокна из обонятельной луковицы. В составе медиального пучка переднего мозга аксоны нейронов септума достигают дорсального таламуса, преоптической области, гипоталамуса. Таким образом, септум является важным связующим звеном между паллиальными формациями и диенцефалоном и входит вместе с медиальным паллиумом в группу лимбических структур мозга.
Вентральная область полушария конечного мозга рептилий представлена несколькими мелкими ядрами, среди которых наиболее полно изучены обонятельный бугорок и nucl. accumbens.
Обонятельный бугорок рептилий получает афферентные проекции
259

Рис. 88. Основные связи стрио-паллидарной системы мозга рептилий
(а) и птиц
(б) (Белехова, 1986).
1 - дорсо-вентрикулярный край, ALa, ALp - ядра лентикулярной петли, DcP – ядро задней комнссуры, DIP - заднее промежуточное ядро таламуса, SpL -спириформное ядро претектальной области.
из обонятельной луковицы (основной), переднего обонятельного ядра, латерального и медиального паллиума. Эфференты бугорка направляются в медиальный паллиум и таламус.
Nucleus accumbens рептилий отличается простотой цитоархи-тектонической и нейронной организации, будучи представлен небольшой группой мелких изодендритных нейронов. Основные афференты он получает от септума, обонятельного бугорка, ДВК, дорсолатерального таламуса, а его эфференты направляются в вентральные районы палеостриатума, преоптическую область, латеральный гипоталамус и вентральную тегментальную область.
Амигдала рептилий представляет собой сложный комплекс ядер, связи и происхождение которых во многом неясны. У рептилий она расположена в вентролатеральной части полушария в задней трети конечного мозга. Выделяется от двух до семи
260
ядер, которые могут объединяться в центральный и базальный комплексы. В состав амигдалы мозга рептилий включают и так называемое сферическое ядро - задний отдел ДВК (оно есть не у всех рептилий и наиболее отчетливо выражено у змей и ящериц-макросматиков). Сферическое ядро получает волокна добавочной обонятельной луковицы и вовлечено в интеграцию обонятельной и соматической информации. Описаны эфферентные связи ядра с преоптической областью и каудальным гипоталамусом. У рептилий помимо филогенетически древних ядер амигдалы, связанных с обонятельной системой (в их число попадает и сферическое ядро), есть новые ядра, связанные с ДВК и корковыми формациями.
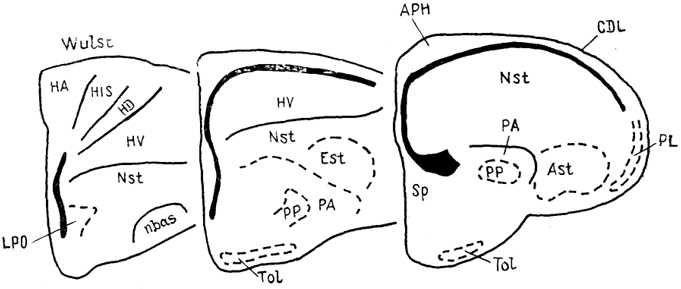
Рис. 89. Строение полушарий мозга голубя
Columba livia на серии фронтальных срезов (Reiner, 1985).
Конечный мозг птиц имеет необычное по сравнению с мозгом других высших позвоночных строение. Латеральные желудочки развиты слабо, почти все полушарие занято диффузными клеточными массами, среди которых выделяют такие структуры, как гипер-, нео-, архи- и палеостриатум (рис. 89). Они разграничены узкими полосками белого вещества.
К собственно паллиальным формациям относят кортикоидную пластинку, расположенную на дорсомедиальной поверхности полушария, и гипер- и неостриатум, которые ранее считали субпаллиальными структурами.
Кортикоидная пластинка мозга птиц максимально развита в каудальных отделах полушария и состоит из дорсомедиального, дорсолатерального отделов и участка латерального паллиума. У многих птиц кортикоидная пластинка имеет сложный нейронный состав и развитую систему связей. Здесь обнаружены пирамидоподобные нейроны и шипиковые и бесшипиковые коротко-аксонные клетки идиодендритного типа. Дорсомедиальный отдел получает афференты от гиперстриатума, дорсолатерального отдела паллиума, архистриатума, гипоталамуса, ряда ядер ствола
261
мозга (ядро шва, голубое пятно) и одноименной структуры из соседнего полушария. Эфференты направляются в септум, дорсолатеральный паллиум, дорсальные отделы гипер- и архистриатума.
Все это дает основание проводить гомологию дорсомедиального отдела паллиума конечного мозга птиц с медиальным паллиумом мозга рептилий и других позвоночных. Структура и связи дорсолатерального отдела кортикоидной пластинки изучены недостаточно.
Латеральный паллиум птиц развит слабо, однако обнаруженные здесь проекции обонятельной луковицы позволяют сравнивать его с латеральным паллиумом рептилий.
Цитоархитектоническая структура гипер- и неостриатума достаточно сложна. Существуют две основные классификации структур полушарий мозга птиц, согласно которым в нем выделяют ряд отделов или полей, степень развития которых сильно варьирует у разных птиц.
Hyperstriatum accessorium (классификация С. Херрика), или поле В (классификация М. Розе) - самый дорсальный отдел гиперстриатума. У большинства изученных видов он разделяется на две части: дорсальную, имеющую кортикоидную структуру и тесно связанную с кортикоидной пластинкой, и вентральную - ядерного типа. Обе части содержат как одиночные нейроны, так и клеточные скопления разного типа. Наиболее значительна дорсальная часть гиперстриатума у пингвинов. Hyperstriatum intercalatus superior (поле А) - наиболее вариабельный по размеру и степени дифференцировки отдел. У одних видов (голуби) он развит слабо, у других (совы, пингвины, вороны) четко выражен. Hyperstriatum dorsale (поле С) есть у всех птиц и отличается наличием большого количества крупных нейронов и нейронных комплексов. Hyperstriatum ventrale (поле D) - самое крупное поле гиперстриатума по размеру как самой структуры, так и составляющих его элементов. Neostriatum (поле G) - структура, простирающаяся до самых каудальных отделов полушария. В его составе выделяют несколько участков: в медиальной части каудального отдела - поле L; в средних отделах - эктостриатум, ectostriatum (поле Е). От соседних участков неостриатума последний отличается однообразием клеточных форм и разделяется на центральную и периферическую части.
Ростральные отделы гиперстриатума образуют особую структуру - дорсальное возвышение, Wulst. По степени его развития и положению выделяется два типа полушарий конечного мозга птиц (рис. 90). Первый характеризуется гипертрофией Wulst и смещением его в ростральные отделы полушария, второй - более слабым развитием и сдвигом в теменно-затылочную область. По первому типу построен конечный мозг голубиных, пингвинов, куриных, чайковых, птиц-дневных хищников, совиных,
262
воробьиных. Интересно отметить, что у перечисленных групп птиц кортикоидная пластинка паллиума развита по-разному. Так, у голубиных, куриных, пингвинов она выражена относительно хорошо, а у воробьиных (особенно у врановых) - плохо. По второму типу построен мозг представителей отрядов ржанковых, пастушковых, голенастых, веслоногих, пластинчатоклювых, а также у дятлов и попугаев. У этих птиц хорошо развита кортикоидная пластинка.
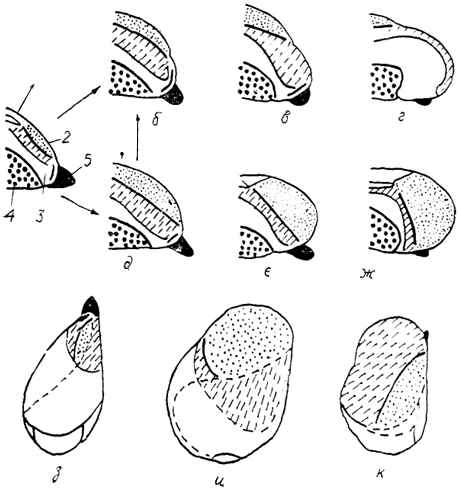
Рис. 90. Два типа организации конечного мозга птиц (Stingelin, 1958).
а - исходный (гипотетический) тип строения полушарий; б-г - полушария второго типа у морского зуйка (б), белого аиста (в) и бекаса (г); д-ж - полушария первого типа у голубя (д), козодоя (е) и грача (ж); з-к - вид мозга сверху: исходный тип (з), у грача (и) и бекаса (к). 1-5 - отделы полушария: 1 - Wulst, 2 - вентральный гиперстриатум, 3 - неостриатум, 4 - палеостриатум, 5 - обонятельная луковица.
Относительный объем полей гиперстриатума сильно варьирует как у представителей разных отрядов, так и в пределах одного отряда или семейства.
Функциональная организация гипер- и неостриатума мозга птиц также очень сложна. В этих структурах обнаружены двойные зоны представительства ряда сенсорных систем. Зрительная информация достигает ростральных отделов НА через дорсолатеральное переднее ядро таламуса, а через круглое ядро - центральной части эктостриатума. Соматосенсорная информация поступает в ростральные отделы НА через дорсальное промежуточное ядро таламуса, а через мультисенсорное дорсолатеральное заднеталамическое ядро - в район на границе каудального
263
неостриатума и вентрального гиперстриатума. Эти две зоны значительно отличаются друг от друга. Нейроны сомато-сенсорной зоны в ростральном гиперстриатуме имеют относительно небольшие рецептивные поля. При этом ростральные отделы тела животного представлены в дорсальной части зоны, каудальные - в вентральной. Нейроны второй соматосенсорной зоны имеют обширные рецептивные поля, получая соматическую информацию от половины, а иногда и всего тела, и являются при этом полимодальными, воспринимая поступающую в соседние районы полушария слуховую и зрительную информацию. Морфологически это обеспечивается длинными дендритами нейронов, выходящими далеко за пределы соматосенсорной зоны. Обе зоны связаны друг с другом двусторонними связями.
В каудальном неостриатуме (поле L) обнаружены входы; слуховой сенсорной системы, волокна которой через овальное ядро таламуса проецируются также в архистриатум, а через медиальную часть дорсолатерального ядра - в ростральные отделы несстриатума.
В свою очередь Wulst и другие поля гипер- и неостриатума образуют обширные связи как внутри полушария, так и с другими отделами мозга. В частности, зрительные зоны Wulst образуют афференты в эктостриатум (поверхностную зону), палеостриатум, дорсальный и вентральный таламус, соседние поля гиперстриатума, претектальную область, тектум. Обнаружены прямые проекции из гиперстриатума в моторные ядра ствола и спинной мозг, аналогичные кортико-спинальным проекциям мозга млекопитающих.
Степень развития полей гиперстриатума (особенно района Wulst) и его нейронных комплексов коррелирует с уровнем сложности поведения птиц. При этом кортикоидная пластинка, гипер- и неостриатум, являясь паллиальными по происхождению, различаются не только по структуре, но и по участию в организации разных поведенческих реакций. Это свидетельствует о высоком уровне дифференцировки паллиальных структур конечного мозга птиц.
У птиц, как и у рептилий, к собственно стриатарным структурам относят палеостриатум и образования вентромедиальной стенки полушария (обонятельный бугорок, септум, nucl. accumbens, часть амигдалы).
В составе палеостриатума различают два поля: палеостриатум возрастающий, paleostriatum augmentatum (поле Н) и палеостриатум примитивный, paleostriatum primitivum (поле J) (рис. 89). Поле РА четко отделяется от вышележащего неостриатума полоской белого вещества и состоит из мелких и средних нейронов, как одиночных, так и образующих скопления (в их состав входит 3-6 нейронов). Поле РР имеет нечеткие границы и находится как бы внутри поля РА. Нейронный состав палеостриатума птиц сходен с таковым у рептилий.
264
Афферентные проекции поле РА получает из зоны РР, неостриатума, гиперстриатума, таламуса, среднего мозга. Многие из этих связей двусторонние. Основные эфферентные связи стри-атума образованы структурами поля PP. Так, обнаружены связи РР с ядром SPL претектальной области, которое связано с тектумом. Таким образом осуществляется связь с моторной системой мозга птиц, поскольку нейроны тектума образуют нисходящие проекции к моторным центрам ствола и спинного мозга.
Стриато-таламо-стриатарные связи осуществляются у птиц через заднее промежуточное ядро, которое получает мозжечковые афференты и проецируется в гиперстриатум, а также через ядра лентикулярной петли. Сформированы двусторонние связи РА и РР с ядром ТРС тегментума среднего мозга, которые гомологичны стриато-нигро-стриатарным связям конечного мозга рептилий и млекопитающих (рис. 88).
Особое место в системе субпаллиальных образований мозга птиц занимают парольфакторная область, lobus parolfactorius, и базальное ядро, nucl. basalis (рис. 89). Парольфакторная область расположена в вентромедиальной части полушария и граничит с полостью мозгового желудочка. Ее часто рассматривают как часть РР, однако ее нейроны не проецируются в РР, образуют связи с ядром ТРС. Скорее всего это самостоятельное-образование субпаллиума птиц.
Базальное ядро расположено в вентральном отделе полушария и имеет обширные связи с различными структурами головного мозга. Оно получает проекции от ядер тройничного нерва и латеральной петли, обонятельного бугорка, периферической части эктостриатума, получающего зрительную информацию. Таким образом, базальное ядро является зоной конвергенции различной сенсорной информации, однако необычно то, что сенсорные проекции из ствола мозга достигают этого отдела полушария, минуя таламус. Исследователи рассматривают данную-структуру как уникальную для птиц и связывают ее функцию с координацией процесса клевания.
Обонятельный бугорок, септум и nucl. accumbens мозга птиц практически не изучены (за исключением общей цитоархитектоники, сходной с таковой у рептилий).
Архистриатум, archistriatum (поле К), конечного мозга птиц, сравнивается с амигдалярным комплексом мозга других позвоночных. Он отчетливо выражен в мозге многих птиц и достигает наибольшего развития у пингвинов. Архистриатум разделяется на базолатеральную и кортикомедиальную части. Довольно крупные радиальные нейроны архистриатума располагаются одиночно. Ростральные отделы образуют связи с таламусом, тектумом, ретикулярной формацией, ядрами ствола; каудальные - с гипоталамусом и гиппокампом.
Таким образом, субпаллиальные отделы конечного мозга птиц имеют черты организации как специфические, так и общие
265
для всех амниот. Это касается формирования двусторонних связей стриатума с паллиальными формациями, с одной стороны, и с диенцефало-мезенцефалическими центрами - с другой.
Конечный мозг млекопитающих обладает наиболее сложными корковыми формациями полушарий. Основу цитоархитектонического деления коры полушарий разработал в начале XX в. К. Бродман. Он разделил кору на две основные области: гомогенетическую, для которой характерно шести-семислойное строение, и гетерогенетическую, где такое строение отсутствует. Гомогенетическая кора разделяется на гомотипическую (шестислойное строение сохраняется и во взрослом состоянии) и гетеротипическую (в процессе эмбриогенеза количество слоев меняется). Каждая из этих территорий разделяется на области, а области на поля, различающиеся по строению, связям, функциональной значимости. Так, у приматов выделено 11 областей, включающих 52 поля.
По классификации И.Н. Филимонова основная область коры, обозначенная им как новая кора (неокортекс, изокортекс) соответствует гомогенетической коре по Бродману. Области старой (архикортекс) и древней (палеокортекс) коры гомологичны только части гетерогенетической коры. Остальную часть коры занимает так называемая межуточная кора (мезокортекс), состоящая из периархикортекса, отделяющего новую кору от старой, и перипалеокортекса, отделяющего новую кору от древней.
В состав новой коры, по мнению И. Н. Филимонова, должен быть включен весь паллиум, в пределах которого стенка мозга уже на самых ранних этапах развития характеризуется наличием основных слоев Гисса (матрикс, промежуточный слой, корковая пластинка, краевой слой). Старая и древняя кора, в отличие от новой, на ранних стадиях развития характеризуется неполнотой строения стенки мозга. Межуточная кора имеет переходный тип строения. К палеокортексу относятся пириформная кора (PC) и препириформная область полушария, а также обонятельная луковица, переднее обонятельное ядро, обонятельный бугорок, септум, часть амигдалы и ряд мелких ядер вентромедиальной стенки полушария.*
В состав архикортекса у млекопитающих включают районы гиппокампа (Н), зубчатой фасции (FD), субикулума (S). Пе-риархикортекс представлен пре- и парасубикулумом (PrS, PaS), taenia tecta (TT), энторинальной (ERC) и цингулярной (СС) корой; перипалеокортекс - переходной инсулярной областью (CI). Исследование большого числа млекопитающих показало, что деление коры на отмеченные выше основные области
266
всюду сохраняет свое значение и проблема их гомологии у представителей разных групп млекопитающих не встречает существенных затруднений.
В эволюции млекопитающих новая кора благодаря своему интенсивному росту оттеснила старую и древнюю кору на медиальную и вентральную поверхности полушария. В результате сформировались две основные борозды: гиппокамповая (sh), отделяющая старую кору от новой, и ринальная (srh), отделяющая древнюю кору от новой. Эти борозды отчетливо выражены в конечном мозге всех млекопитающих. Далее происходило образование борозд и извилнн в пределах корковых областей. По степени развития борозд и извилин полушария мозга млекопитающих разделяются на два типа: лиссэнцефаличеекий, для которого характерно отсутствие или очень слабое развитие борозд на поверхности полушария, и гирэнцефалический - с более или менее развитой системой борозд и извилин. Первый тип характерен для однопроходных, низших сумчатых, насекомоядных, рукокрылых, грызунов. Остальные млекопитающие имеют на поверхности полушария систему борозд и извилин. Вместе с тем необходимо подчеркнуть, что среди разных групп современных млекопитающих есть представители, анатомия мозга которых отличается от характерной для всей группы. Так, у ехидны полушария имеют необычно развитую систему борозд и извилин, захватывающих даже область древней коры (рис. 91).
Межвидовая и межгрупповая изменчивость проявляется в варьировании многих важных показателей уровня развития и дифференцировки коры: площади и объема одноименных областей и полей, толщины коры, степени выраженности и толщины отдельных слоев корковой пластинки, размеров и плотности расположения нейронов.
В процессе эволюции млекопитающих происходило последовательное увеличение площади, занимаемой ассоциативными полями и усиление вариабельности в расположении одноименных зон коры у разных животных (рис. 92).
Каждое поле и каждая область коры имеют многочисленные связи с другими отделами коры и частями мозга. Примерно половина из них образована афферентными волокнами. Они подразделяются на две большие группы: кортико-кортикальные (ассоциативные и межполушарные) и образованные волокнами из нижележащих отделов мозга.
Ассоциативные связи объединяют поля в пределах одного полушария. В образовании этих связей принимают участие нейроны многих слоев коры. Количество, типы и характер распределения по слоям нейронов, образующих ассоциативные проекции, зависят от поля коры, вида животного и, как полагают, могут меняться в процессе онтогенеза. Пирамидные нейроны образуют от 60 до 90% таких связей. Число и распределение
267
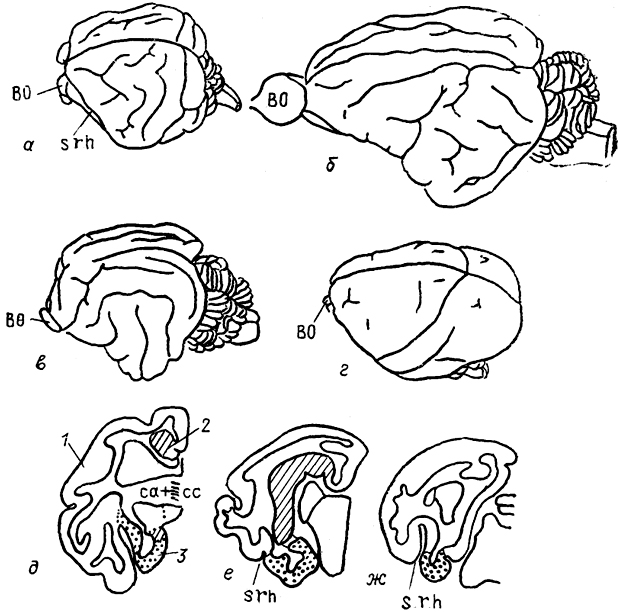
Рис. 91. Внешний вид головного мозга различных млекопитающих (а-г) в поперечные срезы мозга ехидны
(д-
ж) (Welker, Lende, 1980).
а - ехидна, б - грызун капибара, в - кошка, г - беличья обезьяна; 1-3 - отделы коры: неокортекс (1), архикортекс (2), палеокортекс (3).
терминалей ассоциативных волокон зависят от слоя коры, в котором проходят кортико-кортикальные аксоны, а также от типа полей, связанных между собой. Корковые поля образуют в пределах полушария связи с разным количеством полей.
Комиссуральные волокна сосредоточены в основном в мозолистом теле, corpus callosum. Его формирование связано с развитием новой коры, однако при резком увеличении у высших млекопитающих, по сравнению с низшими, площади мозолистого тела, плотность волокон сохраняется на одном уровне: примерно 400 тысяч волокон на 1 мм2 сечения. В коре есть участки, как образующие комиссуральные связи (их большинство), так и лишенные их (поле 17 зрительной коры). Ней-
268
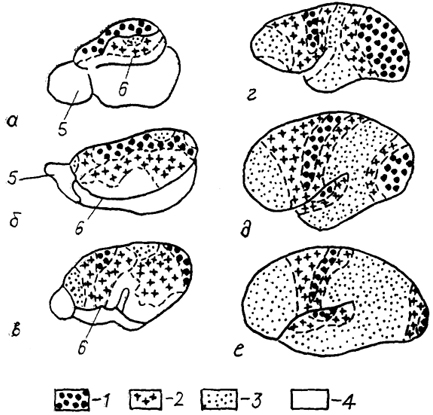
Рис. 92. Расположение и степень развития корковых полей различных типов на латеральной поверхности полушарий млекопитающих (Богословская, Поляков, 1981).
а - еж, б - крыса, в - собака, г - мартышка, д - оранг, е - человек; 1-3 - поля леокортекса: первичные (1), вторичные (2) и третичные (3), 4 - территория древней, старой и межуточной коры, 5 - обонятельная луковица, 6 - ринальная борозда.
роны, образующие каллозальные проекции, как и их окончания, располагаются во всех слоях коры.
Афферентные связи включают кроме перечисленных выше таламо-кортикальные и экстраталамические проекции. У разных животных и в разных полях коры характер окончаний таламических афферентов (даже от одноименных ядер таламуса) может варьировать).
Экстраталамические афференты берут начало в основном от центров ствола и в соответствии с источником проекции разделяются на холин-, адрендофамин-, и серотонинергические. Эти волокна проходят во все слои коры (в каждой системе имеются свои особенности распределения окончаний по слоям), идут параллельно поверхности коры на большом расстоянии (до нескольких миллиметров) и образуют как типичные синаптические соединения, так и модуляторные контакты. Общее число таких окончаний составляет от 5 до 12% от числа межнейронных контактов в коре.
Наиболее крупными эфферентными путями считаются кортико-спинальные (пирамидный тракт) и кортико-бульбарные тракты. Они образованы аксонами пирамидных нейронов нижних (V-VI) слоев коры. Большинство эфферентов (особенно идущих от моторных и сенсорных полей коры) имеют строго топическую
269
организацию. Волокна нисходящих трактов формируют многочисленные коллатерали, оканчивающиеся в различных отделах мозга. Так, коллатерали волокон пирамидного тракта оканчиваются на нейронах красного ядра, образующих руброспинальные проекции, и влияют на их деятельность.
У млекопитающих встречается несколько вариантов строения неокортекса. Первый отмечен у насекомоядных, рукокрылых, грызунов, зайцеобразных. Неокортекс занимает 30-60% общей поверхности коры. Преимущественно развиты нижние слои коры и слой 1. Присутствует большое количество слабодифференцированных, атипичных нейронов. По цитоархитектонике и нейронному составу зоны и поля коры различаются между собой слабо. Среди представителей каждой из этих групп млекопитающих встречаются и крайне специализированные виды, план строения коры которых значительно отличается от такового у всей группы. Так, у ежей обнаружены участки неокортекса, характеризующиеся преимущественным развитием верхних и нижних слоев коры и появлением высокодифференцированных пирамидных и звездчатых нейронов.
Второй тип строения неокортекса встречается у китообразных, конечный мозг которых значительно отличается от мозга других млекопитающих. Он имеет чрезвычайно сложную систему борозд и извилин, неокортекс занимает 97-98% поверхности коры и очень тонкий (0,158-0,202 см), вследствие чего относительный объем неокортекса китообразных оказался меньше, чем у насекомоядных. Цитоархитектоническая и нейронная структура неокортекса имеют многие черты, характерные для мозга низших млекопитающих. При этом в мозге дельфинов обнаружены длинные ассоциативные внутрикорковые связи, развитие которых является специфической чертой конечного мозга приматов. Все это свидетельствует о своеобразном пути эволюции конечного мозга китообразных.
Третий вариант строения неокортекса отмечен у представителей таких отрядов млекопитающих, как хищные, ластоногие, парно- и непарнокопытные, хоботные. В целом их конечный мозг отличается развитием системы крупных дуговых борозд и извилин на латеральной поверхности полушария, сильно затрудняющих сравнение с полушариями конечного мозга других млекопитающих. Неокортекс занимает от 80 до 95% площади коры и имеет большую толщину. Нейронная структура характеризуется увеличением разнообразия и полиморфизма клеток коры, намечается разделение неокортекса на гранулярные и агранулярные поля.
Четвертый тип коры встречается у приматов. В этой группе млекопитающих, филогенетически самой молодой, наиболее высока эволюционная и адаптивная изменчивость. Это проявляется в значительной вариабельности структуры коры в разных полях одного полушария и одноименных полей у разных
270
животных. Резко увеличивается количество и разнообразие клеточных форм в составе неокортекса, завершается выделение гранулярных и агранулярных областей коры, разрастаются височные доли полушария, большую площадь занимают третичные, ассоциативные поля.
В настоящее время созданы различные теории организации неокортекса млекопитающих, среди которых наибольшее распространение получила модульная концепция структурно-функциональной организации коры. Интересную модель эволюции модулей в коре мозга млекопитающих предложил П. Морган.
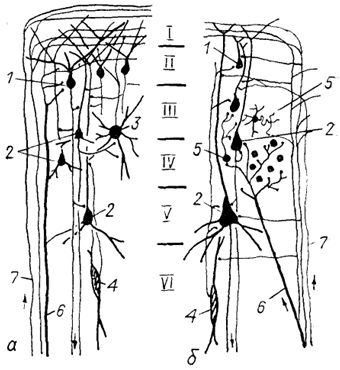
Рис. 93. Схема возможной эволюции модульного строения коры млекопитающих (Glezer e. а., 1988).
а - исходный гипотетический модуль, б - модифицированный модуль. 1 - пирамиды II слоя с широкой зоной ветвления дендритов, 2 - пирамиды III-V слоев, 3 - мультиполярная клетка, 4 - биполярная клетка, 5 - короткоаксонные звездчатые нейроны разных типов, 6 - специфическиеи 7 - неспецифические таламо-кортикальные афференты.
Согласно этой модели в эволюции происходило образование в коре модулей с их последующей модификацией (усложнением) и увеличением количества. Исходный (гипотетический) тип мозга предков млекопитающих характеризовался наличием в: коре небольшого количества просто организованных модулей. В этих модулях специфические и неспецифические таламо-кортикальные волокна, а также ассоциативные и комиссуральные проекции направлялись преимущественно в I слой, контактируя с апикальными дендритам и пирамидных и непирамидных нейронов (рис. 93). Большие мультиполярные изодендритные нейроны посылали свои аксоны во второй слой, обеспечивая внутрикорковые связи. Таким образом, основным "афферентным" слоем коры был II слой с его специализированными пирамидными клетками, аксоны которых направлялись во все нижележащие слои коры.
В процессе эволюции в модулях происходило разделение зон окончаний основных афферентных проекций. Таламо-кортикальные афференты оканчивается преимущественно в новом, интенсивно развивающемся IV слое, где появляется большое число высокоспециализированных звездчатых нейронов. Другие афференты направляются в верхние слои коры, давая на своем протяжении многочисленные коллатерали.
271
В разных группах млекопитающих процесс модификации и увеличения количества модулей шел, по мнению автора этой гипотезы, независимо, в результате чего к настоящему времени сформировалось четыре основных типа неокортекса. Консервативный, при котором сохраняются многие признаки гипотетического типа коры, встречается у представителей современных "отрядов насекомоядных и рукокрылых. Прогрессивно-консервативный тип, характеризующийся некоторым увеличением количества модулей и незначительным их усложнением. Животные с таким типом неокортекса встречаются практически во всех отрядах современных млекопитающих (например, лемуры и мартышки среди приматов). При прогрессивном типе строения •коры количество, степень сложности и разнообразие модулей значительно увеличивается. В каждом отряде млекопитающих есть представители, неокортекс которых имеет черты прогрессивного типа (среди приматов это высшие обезьяны и человек). Консервативно-прогрессивный тип отличается резким увеличением количества модулей в коре при незначительной их модификации, что приводит к формированию "монотонной" структуры коры (китообразные).
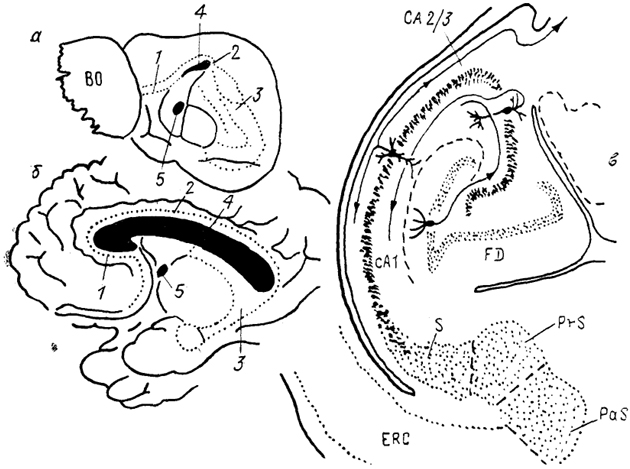
Рис. 94. Организация гиппокампа у ежа
(а) и человека
(б) и строение гиппокампа лемура на тангенциальном срезе
(в) (Stephan, Malonescu, 1980).
1 - прекаллозальный, 2 - супракаллозальный и 3 - посткаллозальный отделы гиппокампа, 4 - мозолистое тело, 5 - передняя комиссура, СА 1 и СА 2/3 - отделы гиппокампа, ERC - энторинальная кора, FD - зубчатая фасция, PaS - парасубикулум, PrS - пресубикилум, S - субикулум.
Старая кора (архикортекс) мозга млекопитающих образована в основном структурами гиппокампа, который разделяется
272
у плацентарных млекопитающих на три отдела: прекомис-суральный (прекаллозальный), супракомиссуральный (супра-каллозальный) и ретрокомиссуральный (посткаллозальный). Из-за сильного развития у высших млекопитающих мозолистого тела первые два отдела у них редуцируются (рис. 94). Прекомиссуральный отдел, расположенный между зоной переднего обонятельного ядра и мозолистым телом, представлен мелкими пирамидными клеткамл и связан афферентными и эфферентными проекциями с пириформной и энторинальной корой. Средняя часть гиппокампа превращается в узкую полоску серого вещества, связывающего передний и задний отделы гиппокампа. Ретрокомиссуральный отдел наиболее развит и представлен структурами субикулума (S) и собственно гиппокампа, который состоит из ряда зон (СА 1-4) и зубчатой фасции (FD). Гиппокамп имеет корковую структуру и состоит из пяти слоев, основным из которых является III слой, образованный пирамидными нейронами. У низших млекопитающих этот слой узкий, компактный; у высших - широкий, диффузный.
Гиппокамп - важнейший компонент лимбической системы конечного мозга, связанный со многими отделами головного мозга. Он получает афферентные проекции от различных зон неокортекса (включая ассоциативные поля), а также септума, амигдалы, ряда ядер таламуса, многих отделов ствола мозга. Эфферентные проекции гиппокампа образованы аксонами исключительно пирамидных нейронов и проходят в основном в составе свода. Через него обеспечивается связь гиппокампа с амигдалой, таламусом, гипоталамусом, палеокортексом, септумом. У высших млекопитающих над мозолистым телом формируется пучок волокон - пояс, cingulum, через который гиппокамп связан с другими отделами архикортекса и рядом полей неокортекса. Комиссуральные связи осуществляются через комиссуру гиппокампа.
Древняя кора (палеокортекс) у млекопитающих занимает в полушарии вентральное положение, оттесненная с латеральной поверхности сильно разросшейся новой корой. Для нее характерны слабая слоистость и неполное отделение от соседних клеточных масс стриатума. Основная часть палеокортекса - пириформная кора - получает обонятельные проекции по латеральному обонятельному тракту, волокна которого оканчиваются на апикальных дендритах пирамидных нейронов пириформной коры. Аксоны этих нейронов направляются в обонятельный бугорок, а также в префронтальную область коры. Палеокортекс имеет эфферентные связи с амигдалой и эпита-ламусом. Обращает на себя внимание тот факт, что у животных, имеющих редуцированную обонятельную систему (например, дельфины, некоторые птицы), палеокортекс развит хорошо. Это свидетельствует о том, что палеокортекс функционально связан не только с обонянием.
273
Субпаллиальные формации, конечного мозга млекопитающих включают в себя комплекс ядер: хвостатое ядро, nucl. caudatus, и скорлупу, putamen, часто рассматриваемые как единая структура (неостриатум), а также бледный шар, globus pallidus, палеостриатум, ограду, claustrum, амигдалу, структуры вёнтромедиальной стенки полушария (септум, обонятельный бугорок, ядро диагональной связки Брока, безымянная субстанция, nucl. accumbens).
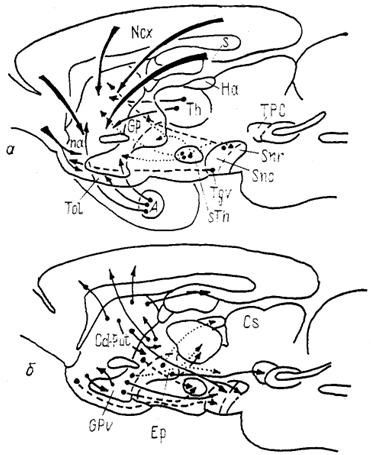
Рис. 95. Основные связи стрио-паллидарной системы млекопитающих на примере мозга крысы (Heimer, Alheid, Zaborzsky, 1985).
а - афферентные, б - внутренние и эфферентные связи.
Цитоархитектоническая и нейронная организация стриатума млекопитающих (он включает формации нео-, палеостриатума и ограду) очень сложна. Неостриатум цитоархитектонически неоднороден. Его клетки образуют сложные пространственные комплексы-стриосомы, окруженные более рыхло расположенными нейронами, составляющими так называемый матрикс. Выделено несколько типов нейронов, различающихся по размеру (крупные, средние, мелкие), по типу аксонов (длинно- и короткоаксонные) и характеру дендритных ветвлений (густо- и редковетвистые; шипиковые и бесшипиковые). Показано, что клетки любого размера могут быть как длинноксонными, образующими эфферентные проекции, так и короткоаксонными, фомирующие внутристриарные связи. Крупные длинноаксонные нейроны дают восходящие проекции в кору, тогда как мелкие и средние длинноаксонные нейроны - нисходящие проекции. В эволюции млекопитающих доля мелких нейронов разного типа увеличивается.
Бледный шар разделяется цитоархитектонически на внутренний и наружный сегменты, нейронный состав которых довольно однообразен. Это в основном крупные редковетвистые нейроны ретикулярного типа со строго упорядоченной ориентацией дендритов,
274
образующих систему микромодулей в виде дисков, параллельных друг другу и латеральной границе ядра, но расположенных перпендикулярно направлению хода аксонов нейронов неостриатума. Таким образом, аксоны клеток хвостатого ядра и скорлупы, проходя в бледный шар, пронизывают сотни дендритных модулей и образуют сложные синаптические комплексы.
Стриатум млекопитающих имеет сложную систему связей с большим числом мозговых структур (рис. 95). Основной тенденцией в организации афферентных и эфферентных проекций стриатума является формирование кольцевых связей: неокортекс → неостриатум → неокортекс, неокортекс → неостриатум → палеостриатум → таламус → неокортекс, неостриатум → палеостриатум → неостриатум, неостриатум → палеостриатум → таламус → неостриатум, стриатум → черная субстанция → стриатум. Многие из этих связей топографически упорядочены. Например, в случае корково-неостриатарных связей нейроны глубоких слоев коры (V, VI) из определенной зоны неокортекса проецируются в стриосомы, верхних (II, III) - в матрикс. При этом входы из различных полей коры в неостриатуме могут перекрываться, что создает основу для конвергенции на нейронах стриатума разнообразной информации.
Таким образом, филогенетически молодая система связей стриатума с корковыми формациями получает у млекопитающих дальнейшее развитие, обеспечивая ему роль одного из интегративных центров конечного мозга.
Ограда - тонкая полоска серого вещества, отделенная от скорлупы пучками волокон, образующих наружную капсулу, capsula externa, а от коры ее отделяет узкая полоска белого вещества - самая наружная капсула, capsula extrema. Для нейронного строения ограды характерна значительная полиморфность. Наиболее многочисленны корковые выходы из различных зон неокортекса. Афференты также поступают из стриатума, амигдалы, таламуса и черной субстанции. Эфференты ограды направляются к новой коре, стриатуму, амигдале, обонятельной луковице, таламусу и мозжечку. Есть данные, что ограда имеет корковое происхождение.
В септальной зоне конечного мозга млекопитающих, включающей медиальное и латеральное септальные ядра, ядро диагональной связки Брока, nucl. accumbens, и функционально связанной с лимбической системой, появляется, в отличие от аналогичной зоны конечного мозга рептилий и птиц, большое количество высокоспециализированных короткоаксонных нейронов. Однако, анализ связей, структуры и гистохимических характеристик позволяют проводить гомологию основных отделов септальной зоны мозга рептилий, птиц и млекопитающих.
Обонятельный бугорок имеет слоистое строение (различают наружный слой волокон, средний слой пирамидоподобных нейронов
275
и внутренний слой полиморфных клеток). Степень выраженности слоев у разных видов млекопитающих значительно варьирует. Афферентные проекции поступают из обонятельной луковицы, пириформной коры, амигдалы, преоптической области, ряда зон неокортекса, интраламинарных ядер таламуса, черной субстанции. Эфференты направляются в септальную зону и далее в гиппокамп, стриатум, бледный шар, дорсомедиальную область таламуса и эпиталамус, амигдалу и пириформную кору. Таким образом видно, что обонятельный бугорок млекопитающих связан не только с обонятельной, но и с лимбической и стриопаллидарной системами.
Амигдала млекопитающих состоит из большого числа (до 10) ядер, которые объединяются в несколько групп: базолатеральную (латеральное, основное и добавочное ядра) - наиболее филогенетически молодую часть амигдалы, связанную с корковыми формациями; кортикомедиальную (периамигдаляр-ная кора, среднее и заднее кортикальные ядра, медиальное ядро) - филогенетически более старую часть, связанную с обонятельной системой; центральную (центральное ядро, переднеамигдалярная зона) - связанную с гипоталамусом и ядрами ствола мозга, контролирующими вегетативные функции организма. В филогенезе млекопитающих в амигдале происходило два процесса. Во-первых, уменьшалась доля, занимаемая ядрами кортикомедиальной группы, и увеличивался объем базолатеральной группы ядер (последняя достигает наибольшего развития и максимальной сложности у приматов и человека). Во-вторых, у высших млекопитающих вследствие сильного развития новой коры амгидала смещается с вентральной стенки полушария на вентромедиальную. Многие вопросы организации амигдалярного комплекса конечного мозга высших позвоночных еще не решены.
Основной вопрос, который возникает при изучении конечного мозга, касается гомологии его структурных подразделений у представителей разных групп позвоночных.
В паллиальных отделах конечного мозга медиальный паллиум и часть дорсального паллиума мозга рептилий рассматриваются как гомологи медиодорсальной кортикоидной пластинки птиц и структур архикортекса млекопитающих; ДВК и часть дорсальной коры рептилий - гипер- и неостриатуму птиц и неокортексу млекопитающих; латеральный паллиум рептилий сравнивается с дорсолатеральным паллиумом птиц и пириформной корой млекопитающих. В субпаллиуме возможными гомологами считаются такие структуры как: неостриатум рептилий - палеостриатум возрастающий птиц - стриопаллидарный комплекс (хвостатое ядро и скорлупа) млекопитающих, а также палеостриатум рептилий - часть палеостриатума примитивного птиц и бледный шар млекопитающих.
Проведение сравнений между более мелкими подразделениями
276
крайне затруднительно, поскольку развитие паллиума и субпаллиума (и их отдельных образований) шло в разных линиях высших позвоночных независимо и параллельно, благодаря чему гомологичные отделы могли приобрести разную структурную организацию.
277
*
В настоящее время исследователи на основании гистохимических, ходологических и эмбриологических данных относят перечисленные здесь структуры (начиная с обонятельного ядра) к субпаллиальным формациям.
