У круглоротых морфологически выражены все основные отделы конечного мозга. В паллиуме полушария у миног описывают четыре отдела: медиальный, субмедиальный (субгиппокампальная доля), дорсальный и латеральный. Субпаллиум, представленный структурами переднего обонятельного ядра, стриатума и септума, у миног дифференцирован слабо. В срединной части каудального отдела полушария расположено преоптическое ядро (рис. 83).
Наиболее дифференцированным отделом полушария миног является медиальный, клетки которого имеют развитую систему дендритных ветвлений и образуют слоистую структуру. Субмедиальный, дорсальный и латеральный отделы паллиума значительно уступают по уровню нейронной дифференцировки медиальному и состоят из мелких нейронов изодендритного типа. Основная масса клеток сосредоточена вблизи латерального желудочка.
У миксин конечный мозг значительно отличается от такового миног. Это относительно крупный отдел, в котором полушария
243
трудно отделить от каудальной части конечного мозга из-за редукции, по-видимому, вторичной, латеральных желудочков. В полушариях выделяют лежащий медио-дорсально медиальный паллиум и латеральную часть, которую трудно сравнивать с какими-либо конкретными структурами паллиума мозга миног из-за необычного строения (в ней выделяют до шести слоев нейронов и их отростков). Субпаллиум миксин занимает в
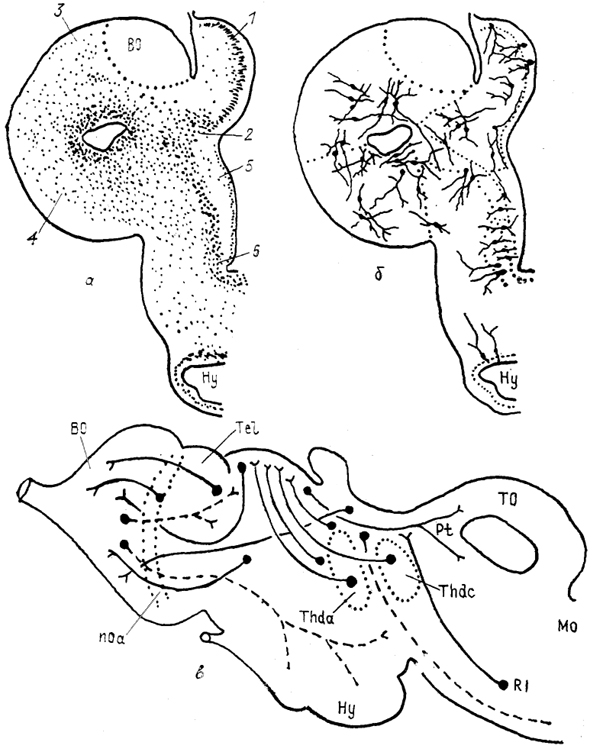
Рис. 83. Схемы строения
(а), нейронной организации
(б) и проводящих путей
(в) конечного мозга миноги
Lampetra fluviatilis (Haier, 1947; Поленова, 1990).
1 - примордиальный гиппокамп, 2 - субгиппокампальная доля, 3 - дорсальный и 4 - латеральный паллиум, 5 - стриатум, 6 - преоптическая зона.
244
полушарии медиовентральное положение и по структуре сходен с субпаллиумом миног. Система афферентных связей конечного мозга круглоротых представлена в основном обонятельными трактами, которые проецируются у миног билатерально в дорсальный и латеральный паллиум, а также контрлатерально в переднее обонятельное ядро и преоптическую область. Субгиппокампальная доля, вероятно, не получает прямых обонятельных проекций. Другие афференты конечного мозга происходят
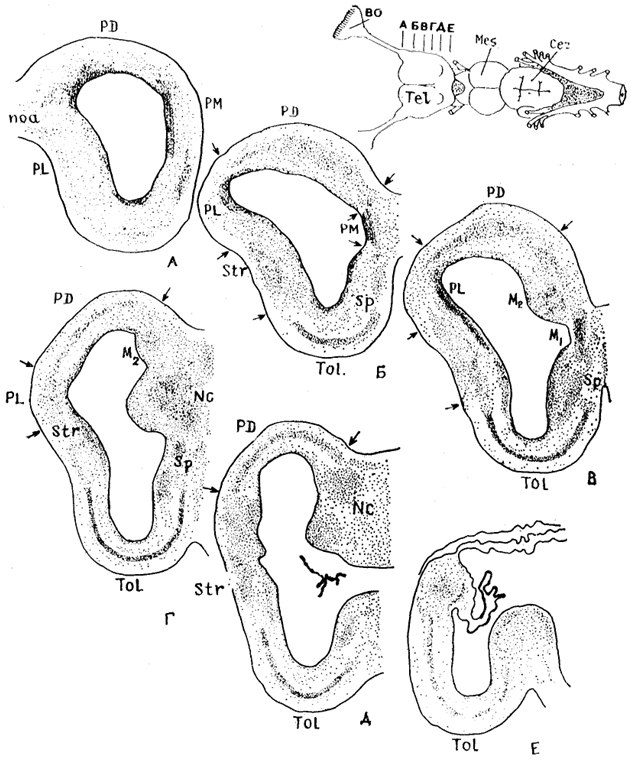
Рис. 84. Строение конечного мозга акулы
Squalus acanthias (Обухов, 1987).
А-Е - уровни срезов мозга; Ml - медиальный и М2 - дорсомедиальный участки медиального паллиума, NC - центральные ядра. Стрелки - границы зон мозга.
245
из ядер дорсального таламуса и проводят информацию от зрительной и соматосенсорной систем (рис. 83). Они оканчиваются в областях полушария, как получающих обонятельные входы, так и свободных от них.
Эфференты паллиума осуществляют центрифугальную иннервацию обонятельной луковицы, кроме того, медиальный паллкум связан с преоптической областью, гипоталамусом и претектальной областью среднего мозга.
Таким образом, у круглоротых сформированы все основные отделы конечного мозга, характерные для других позвоночных, которые получают проекции от структур обонятельной, зрительной, ссматосенсорной систем. Медиальный отдел паллиума мозга круглоротых сравнивается с медиальным (гиппокампальным) паллиумом мозга челюстноротых позвоночных, субгиппокампальный с дорсальным, а дорсальный и латеральный отделы - с латеральным пириформным паллиумом. Вместе с тем современные круглоротые являются продуктом длительной эволюции, проходившей во многом независимо и параллельно эволюции в линии челюстноротых позвоночных, что не позволяет считать их мозг подобным мозгу предков позвоночных.
В обширном надклассе Рыбы встречаются самые различные варианты строения конечного мозга: от инвертированного у хрящевых и двоякодышащих, до эвертированного у лучеперых.
Структура полушарий у изученных хрящевых рыб чрезвычайно разнообразна. У многих акул и скатов парность полушарий внешне выражена только в самой ростральной части мозга (см. рис. 5). Выделяются виды, полушария которых имеют хорошо развитые латеральные желудочки (например, у акулы Squilus acanthias), и виды, у которых они практически редуцированы (например, у акулы Mustelus canis или ската Raja clavata).
Цитоархитектоника и состав конечного мозга подробно изучены только у некоторых представителей хрящевых рыб (в основном пластиножаберных). В паллиуме конечного мозга акулы Squalus acanthias выделяют три отдела: медиальный, дорсальный и латеральный, которые гомологичны соответствующим отделам паллиума конечного мозга инвертированного типа других позвоночных (рис. 84). У данного вида акул наиболее дифференцированным отделом паллиума является дорсальный, в пределах которого, впервые у позвоночных, формируется корковая пластинка - слой клеток, отделенных от перивентрикулярной зоны. В ней обнаружены высокодифференцированные пирамидоподобные и звездчатые короткоаксонные нейроны, сходные по структуре с аналогичными клеточными элементами корковой пластинки паллиума высших позвоночных.
В каудальной части конечного мозга дорсальный отдел полушария представлен так называемым центральным ядром. Медиальный
246
паллиум занимает большую часть дорсомедиальной стенки полушария и разделяется на две зоны: медиальную (M1) и дорсомедиальную (М2). Медиальная зона наиболее выражена в ростральных отделах полушария, дорсомедиальная - в средних и каудальных отделах полушария, отличается ют зоны M1 присутствием крупных нейронов радиального типа.
В латеральном паллиуме нейроны мигрируют в толщу стенки полушария, однако в отличие от дорсального паллиума не формируют корковой пластинки. Эта часть полушария представлена популяциями изодендритных нейронов лофодендритного и радиального типов. У некоторых хрящевых рыб, в частности у акулы Squalus acanthias, в каудальных отделах латерального паллиума, на границе со стриатумом, выделяют группу нейронов - ядро А, которое рассматривается как гомолог паллиальной части амигдалы.
Субпаллиальные отделы конечного мозга хрящевых рыб значительно развиты и представлены стриатумом, обонятельным бугорком, септумом и амигдалой (рис. 84). У многих изученных акул стриатум разделяется на дорсальный и вентральный отделы, которые отличаются сложным нейронным составом и по структуре и гистохимическим особенностям (связи изучены недостаточно) сравниваются с компонентами стрио-паллидарной системы высших позвоночных. Септум занимает медиовентральную часть стенки полушария и разделяется на медиальное и латеральное ядра.
У скатов, среди которых наиболее изучены представители отрада Rajaformes, структура полушарий резко отличается от таковой у многих акул. Так, латеральные желудочки практически редуцированы, а паллиум и субпаллиум представлены диффузными клеточными массами, среди которых выделить какие-либо отделы крайне трудно.
Интересно отметить, что на ранних стадиях развития нервной системы ската Raja clavata в полушариях конечного мозга формируются зачатки паллиальных формаций и есть отчетливо выраженные желудочки. Только к 71-73 суткам развития в паллиуме и субпаллиуме начинается активная миграция клеток и полушария приобретают свойственную взрослым формам диффузную структуру. Учитывая то, что скаты, как полагают, произошли в эволюции от акулоподобных предков, данные преобразования структуры конечного мозга можно рассматривать как вторичные.
Исследование афферентных связей конечного мозга пластиножаберных рыб показало, что зона прямых обонятельных проекций ограничена рядом отделов полушария (переднее обонятельное ядро, латеральный паллиум, обонятельный бугорок, часть стриатума, септум). В корковом слое дорсального паллиума зарегистрированы ответы на световые и обонятельные воздействия, центральное же ядро является местом окончания таламических
247
афферентов, приносящих в полушарие информацию о зрительных раздражителях. Эти проекции топографически обособлены: вентролатеральное ядро таламуса, связанное с сетчаткой, проецируется в среднюю часть центрального ядра; латеральное ядро, связанное с тектумом, проецируется в каудальные отделы. Это свидетельствует о формировании у некоторых хрящевых рыб двух самостоятельных систем связей: ретино-таламо-теленцефальной и ретино-текто-таламо-теленцефальной. В центральное ядро, а также в обонятельный бугорок поступают афференты от таламических ядер, связанных с мозжечком и органами боковой линии.
Эфференты изучены недостаточно подробно. Они берут начало как в паллиальных, так и в субпаллиальных отделах полушария и адресованы обонятельным луковицам, таламусу, нижней доле гипоталамуса, стволу и верхним сегментам спинного мозга.
Основная масса проекций проходит в составе двух крупных трактов: медиального и латерального пучков переднего мозга. Комиссуральные волокна идут в основном в составе передней и задней комиссур конечного мозга, расположенных в средних иг каудальных отделах полушарий.
В целом, среди современных хрящевых рыб имеется значительная вариабельность в организации одноименных структур конечного мозга, а у некоторых акул уровень его структурно-функциональной организации по ряду параметров (структурных, физиологических, биохимических) сравним с таковым у высших позвоночных.
Структура конечного мозга двоякодышащих и кистеперых рыб изучена далеко неполно. Полушария развиваются путем инверсии, однако их организация у представителей двух основных групп Dipnoi значительно различается. У Neoceratodus forsteri практически отсутствует медиальная стенка полушария, а на ее месте в ростральных отделах мозга развивается мембранозная оболочка, septum ependimale, лишенная нейронов, тогда как у Protopterus dotloi медиальная стенка хорошо выражена, и полушария имеют типичное строение. Несмотря на то, что паллиум двоякодышащих рыб развит относительно слабо и уступает по размеру субпаллиуму, в его дорсальном отделе формируется, как и у некоторых хрящевых рыб, корковая пластинка. Среди субпаллиальных структур выделяют стриатум, септум, обонятельный бугорок, нейронный состав и связи которых практически не изучены.
Инвертированный мозг единственного в современной фауне представителя кистеперых рыб - латимерии - имеет необычное строение: в ростральных отделах конечного мозга дорсолатеральная стенка полушария сильно разрастается и представлена диффузной массой клеток. В средних и каудальных отделах в паллиуме выделяют медиальный, дорсальный и латеральный
248
участки, клетки которых сосредоточены в основном в перивентрикулярной зоне. Отделы субпаллиума (стриатум, септум, обонятельный бугорок) дифференцированы слабо.
Конечный мозг лучеперых рыб в силу особенностей его формирования в эмбриогенезе значительно отличается от такового других позвоночных (см. рис. 4). В эвертированных полушариях выделяют две основные области: дорсальную (D) и вентральную (V), разделяемые в свою очередь на зоны.* У представителей различных групп лучеперых рыб степень дифференцировки полушарий значительно варьирует, однако четко прослеживается эволюционная тенденция усложнения их структуры у представителей высших отрядов по сравнению с низшими (рис. 85). Так, у многопера Polypterus ornatipinnis дорсальный отдел полушария не дифференцирован на зоны, а основная масса нейронов сосредоточена в узком субэпендимном слое под поверхностью полушария. Вентральная область разделяется на три зоны: дорсальную (Vd), вентральную (Vv) и латеральную (VI). У хондростей и голостей, наиболее изученными представителями которых являются осетровые и ильные рыбы, в дорсальной области уже дифференцируются три или четыре зоны: медиальная (Dm), дорсолатеральная (Dd+l), разделяемая часто на дорсальную (Dd) и латеральную (D1), и центральная (Dc). Нейроны активно мигрируют в толщу стенки полушария, формируя в ряде случаев (осетры) слоистые структуры.
У костистых рыб встречаются различные варианты строения паллиума и субпаллиума: от слабодифференцированных у низших, до сложнейшего комплекса у представителей высших костистых рыб, где в составе полушария выделяют до полутора десятков отдельных зон.
Характер связей конечного мозга лучеперых рыб изучен далеко не полностью. Однако совершенно очевидно, что, с одной стороны, зона окончания обонятельных афферентов охватывает не весь конечный мозг, а с другой - ряд областей и зон полушария, как получающих обонятельные входы, так и лишенных их, имеют проекции других сенсорных систем. Так, у костистой рыбы Sebasticus marmoratus обонятельные афференты проецируются билатерально в каудальный отдел дорсальной области - зону Dp, часть зоны DC и ряд районов вентральной области (зоны VI, Vi, Vs). В зоны Dm и Dd приходят афференты из преоптической области и ядер вентрального таламуса, вероятно связанного с акустико-латеральной системой. Зона D1 получает волокна от ядер таламуса, связанных с тектумом. Из стволовой части мозга (в частности, из ядер шва и голубого пятна) афферентные волокна билатерально достигают зон Dc, Dd и дорсальной части медиальной зоны.
249
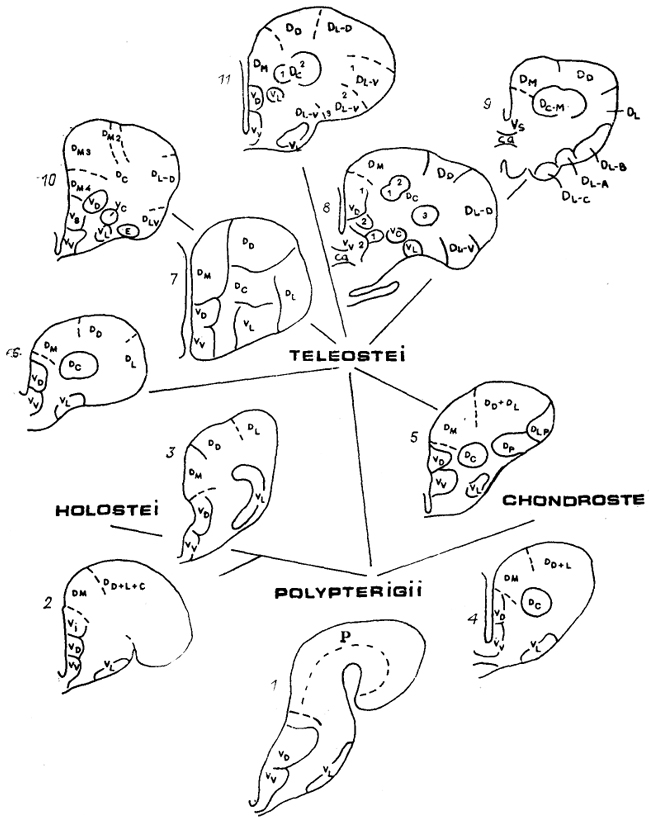
Рис. 85. Схема строения полушарий конечного мозга разных лучеперых рыб: многоперы
(1), грлостеи
(2, 3), хондростеи
(4, 5), низшие
(6, 7) и высшие
8-11) костистые.
Буквами обозначены зоны полушарий по классификации R. Nieuwenhuys.
Эфферентные связи полушария, проходящие в основном по латеральному пучку переднего мозга, образованы аксонами нейронов зон DC, Dd и Dm. Они проецируются ипсилатерально в таламус, нижнюю долю гипоталамуса, средний мозг. Через медиальный обонятельный тракт центрифугальные волокна из зон Dp, DC, Vd достигают обонятельной луковицы.
250
Внутренние связи полушарий обеспечивают взаимодействие между их областями и зонами. Так, зона DC ипсилатерально связана с соседними зонами паллиума, а через переднюю комиссуру с зоной D1 соседнего полушария. Отмечены двусторонние связи медиальной зоны. Дорсальная зона паллиума посылает .многочисленные волокна в медиальную зону, откуда берут начало эфференты к нижней доле гипоталамуса.
Сходные результаты получены при изучении связей конечно-то мозга и других представителей лучеперых рыб.
В связи с особенностями эмбриогенеза конечного мозга лучеперых рыб сложным является вопрос гомологии их полушарий со структурами конечного мозга инвертированного типа. Одни исследователи предлагают проводить прямую гомологию зон :мозга двух типов, другие считают невозможным проведение каких-либо сравнений, поскольку рассматривают конечный мозг .лучеперых рыб как особую линию в эволюции мозга позвоночных. Третьи, а их большинство, основываясь на современных данных о морфологии, гистохимии и связях конечного мозга позвоночных и учитывая особенности процессов инверсии и эверсии, считают, что можно найти решение данной проблемы. Так, зона Dp рассматривается ими как возможный гомолог латерального паллиума, зоны D1 и Dd - медиального и дорсального паллиума соответственно. Каудальные отделы зоны Dm и часть зоны DC сравнивают со стриатумом, отмечая при этом, что другие части этих зон включают компоненты дорсального паллиума. Ростральные отделы медиальной зоны гомологизируются с частью амигдалы. Вентральные зоны Vd и Vv сравниваются с областью септальных ядер, зона VI - с обонятельным бугорком, а каудальные отделы Vs, Vp, Vi- с частью амигдалы (рис. 86).
В целом конечный мозг лучеперых рыб отличает высокий уровень структурно-функциональной организации. Он получает проекции различных сенсорных систем (обонятельной, зрительной, соматической), а через систему эфферентных связей принимает участие в регуляции многих физиологических реакций рыб. Вместе с тем, несмотря на попытки провести гомологизацию отделов конечного мозга лучеперых рыб и других позвоночных, задача эта однозначно не решена.
У амфибий в паллиуме выделяют медиальный, дорсальный и латеральный отделы. У бесхвостых амфибий паллиум отделяется от субпаллиума полосками белого вещества, у хвостатых и безногих эта граница выражена только в районе медиального паллиума.
У всех амфибий наиболее дифференцирован медиальный паллиум, где нейроны активно мигрируют в толщу стенки мозга, тогда как в других отделах они сосредоточены в основном в пе-ривентрикулярной зоне. У высших бесхвостых амфибий (жаб, жерлянок, квакш) в медиальном паллиуме цитоархитектонически выделяют две зоны: медиальную и дорсомедиальную.
251
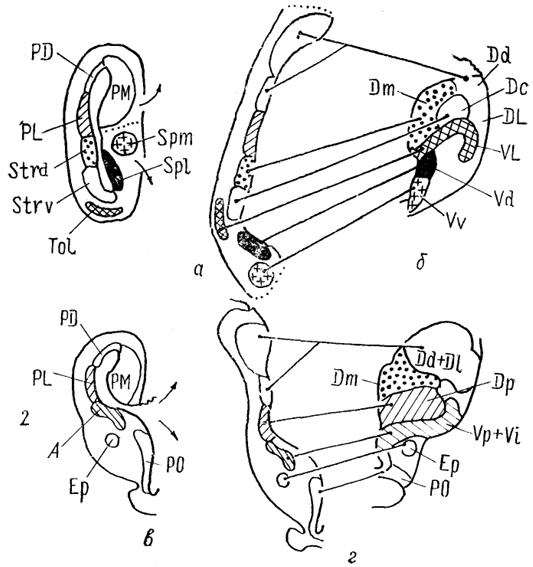
Рис. 86. Схема возможных гомологии структур инвертированного
(а, в) и эвертированного
(б, г) конечного мозга (Northcutt, 1981).
1 - ростральные и 2 - каудальные отделы полушарий.
Большинство данных, касающихся проводящих систем конечного мозга, получено на представителях бесхвостых амфибий, хвостатые и безногие в этом отношении менее изучены.
Афферентные проекции паллиума направляются главным образом в его медиальный отдел. По медиальному обонятельному тракту сюда поступает обонятельная информация, а по медиальному пучку переднего мозга через переднее ядро дорсального таламуса медиальный паллиум получает афференты зрительной, слуховой и соматосенсорной систем, а также гипоталамические проекции.
Эфференты медиального паллиума направляются в соседние зоны паллиума, септум, амигдалу (медиальную часть), вентральный таламус и гипоталамус и организованы в пучки, аналогичные своду, fornix, конечного мозга высших позвоночных. Отделы медиального паллиума полушарий соединены друг с другом паллиальной комиссурой - аналогом комиссуры гиппокампа высших позвоночных.
Дорсальный паллиум имеет обширные двусторонние связи с
252
соседними зонами паллиума, прямые обонятельные и таламические проекции отсутствуют. Латеральный паллиум является основной областью окончаний обонятельных проекций, достигающих его по латеральному обонятельному тракту. Эфференты широко распределяются среди паллиальных и субпаллиальных образований, а также направляются в ипсилатеральную обонятельную луковицу.
Субпаллиальные отделы конечного мозга амфибий включают стриатум, септум, обонятельный бугорок, амигдалу (медиальную часть), переднее обонятельное ядро и nucl. accumbens.
Стриатум - самый большой отдел субпаллиума амфибий - образован мелкими изодендритными нейронами, сосредоточенными в перивентрикулярной зоне. У бесхвостых амфибий стриатум разделяется на дорсальный и вентральный отделы, различающиеся плотностью расположения клеток. Основные проводящие пути стриатума проходят по системе латерального пучка переднего мозга. В таламусе источниками афферентных проекций в стриатум являются ядра центрального и латерального отделов. Одно из этих ядер (латеральное) связано с тектумом и передает в стриатум зрительную информацию. Незначительные афферентные проекции стриатум получает из вентрального гипоталамуса, преоптической области, латеральной части амигдалы и энтопедункулярного ядра. Особо следует отметить формирование у амфибий двусторонних связей стриатума с претектальной областью и тегментумом, что позволяет сравнивать их со стриато-нигро-стриатарными связями мозга высших позвоночных. С паллиумом стриатум амфибий не связан.
Септум располагается на медиовентральной стенке полушария и разделяется на два ядра: медиальное и латеральное. Степень их развития у представителей разных отрядов амфибий различна. У хвостатых (например, у Triturus vulgaris медиальное ядро развито слабо и практически не отделено от латерального, а нейроны обоих ядер относятся к разновидностям клеток изодендритного типа. У бесхвостых амфибий (Rana temporaria, Bufo bufo) медиальное ядро четко отделяется от латерального, а в их составе появляются многочисленные радиальные и биполярные нейроны (дендриты последних связывают два септальных ядра). По медиальному пучку переднего мозга в септум поступает соматосенсорная и зрительная информация из таламуса, а по медиальному обонятельному тракту - обонятельная (в ростральные отделы септума). Эфференты направляются в преоптическую область, вентральный таламус и гипоталамус. Септум имеет также обширные двусторонние связи с медиальным и дорсальным паллиумом.
Обонятельный бугорок достаточно подробно описан лишь в мозге бесхвостых амфибий, где он отчетливо выявляется в вентральных отделах полушария и представлен скоплением мелких слабодифференцированных нейронов. Основные афференты
253
поступают к нему по медиальному обонятельному тракту от ипси- и контрлатеральной обонятельной луковицы, а также от медиального паллиума. Эфференты практически не изучены; (они отмечены только в медиальном паллиуме). Этот отдел мозга амфибий часто сравнивают с разными структурами конечного мозга высших позвоночных (обонятельным бугорком, ядром диагональной связки Брока, частью медиального септального ядра).
Амигдала амфибий занимает в каудальных отделах полушария значительную часть вентролатеральной стенки мозга и разделяется иа два участка: медиальный (субпаллиальный) и латеральный (паллиальный). Медиальная амигдала имеет довольно сложный нейронный состав и формирует в основном двусторонние связи с медиальным паллиумом, стриатумом, гипоталамусом и таламусом. Латеральная амигдала представлена группой мелких веретеновидных нейронов, расположенных вблизи латерального желудочка, и является единственной структурой полушария, получающей проекции от добавочной обонятельной луковицы. Эфференты достигают гипоталамуса, вентрального таламуса, тектума.
Многие исследователи ранее рассматривали мозг современных амфибий как структуру, которая претерпела в процессе-эволюции определенное упрощение. Об этом свидетельствуют, в частности, перивентрикулярный характер расположения клеток в большинстве отделов паллиума и субпаллиума, отсутствие во взрослом состоянии корковой пластинки, большое количество изодендритных нейронов. Вместе с тем, в конечном мозге современных амфибий (особенно бесхвостых) есть признаки, говорящие о достаточно высоком уровне его структурно-функциональной организации. Так, основная и добавочная обонятельные луковицы имеют раздельные области представительства в полушариях (латеральный паллиум и латеральная часть амигдалы соответственно). Формируются две области конвергенции разно-модальной сенсорной информации, поступающей в полушарие - в медиальном паллиуме и стриатуме. Полагают, что у амфибий в медиальном паллиуме закладывается система связей, аналогичная таковой в гиппокампе мозга высших позвоночных и являющаяся важным компонентом лимбической системы. Стриатум участвует в формировании связей между основными уровнями экстрапирамидной системы - теленцефальным и среднемоз-говым. Все это позволяет конечному мозгу принимать участие в регуляции важнейших функций организма амфибий.
В целом для конечного мозга низших позвоночных характерны различные варианты организации: это и высокодифференцированный мозг некоторых хрящевых рыб, необычный по структуре мозг кистеперых рыб и эвертированный мозг лучеперых рыб, что свидетельствует о независимой и параллельной эволюции конечного мозга в разных группах анамний. При этом
254
базовой чертой организации их конечного мозга является наличие в его структурах зон представительства разных сенсорных систем (обонятельной, зрительной, слуховой, соматической) и, как следствие этого, выполнение конечным мозгом функций одного из интегрирующих центров головного мозга.
255
*
Вентральная область полушарий конечного мозга лучеперых рыб не подвергается эверсии и ее положение сходно с таковым вентральной (субпаллиальной) области мозга инвертированного типа.
