Конечный мозг, Telencephalon, анатомически разделяется на две части: каудальную (непарную), telencephalon impar, непосредственно переходящую в структуры промежуточного мозга, и ростральную, образованную парными полушариями, hemis-pherae telencephali. Все отделы конечного мозга развиваются в юнтогенезе как производные крыловидной пластинки. При этом уже на ранних стадиях формирования переднемозгового пузыря, матричная зона, где сосредоточены камбиальные элементы и откуда начинается активная миграция нейробластов, образующих :в дальнейшем те или иные структуры полушария, разделяется на две зоны: дорсальную и вентральную. Производными дорсальной матричной зоны являются формации паллиума, вентральной - субпаллиума.
В разных группах позвоночных процессы развития паллиальных формаций могут протекать по-разному, в результате чего образуется звертированный или инвертированный тип полушарий и конечного мозга в целом (см. рис. 4). Эвертированный тип полушарий встречается только у лучеперых рыб, а у остальных позвоночных они инвертированы. Субпаллиум процессу эверсии ни у кого из позвоночных не подвергается.
В ростральных отделах полушарий происходит образование ларных выпячиваний стенки мозга и формирование обонятельных луковиц.
Обонятельные луковицы, bulbus olfactorius, есть у большинства позвоночных (случаи их редукции у некоторых позвоночных, например ряда птиц и китообразных, рассматриваются эволюционно как вторичные). У круглоротых, большинства костистых рыб, амфибий, черепах, птиц и многих млекопитающих обонятельные луковицы непосредственно примыкают к основной части полушарий, тогда как у хрящевых, двоякодышащих, некоторых костистых рыб, ряда амфибий, большинства рептилий и у приматов они отнесены от нее на некоторое расстояние с помощью так называемых обонятельных ножек, по которым
240
проходят обонятельные тракты. Обычно обонятельные луковицы расположены относительно основной части полушарий рострально или ростро-вентрально, и только у акул лежат латерально (см. рис. 5). Их форма варьирует от овальной до конусовидной.
Обонятельная луковица у всех позвоночных имеет сходный план строения и состоит из нескольких концентрически расположенных клеточных и волокнистых слоев, структура которых значительно варьирует (рис. 82).
Волокна обонятельного нерва, войдя в луковицу, разветвляются и участвуют в формировании особых структур - клубочков, или гломерул, где они контактируют с дендритами различных нейронов луковицы. У низших позвоночных каждое волокно может участвовать в образовании нескольких клубочков, которые располагаются на разной глубине. В связи с этим клубочковый слой выражен неотчетливо. У высших позвоночных каждое волокно обонятельного нерва образует только одну гломерулу, а их слой компактный и ясно выражен.
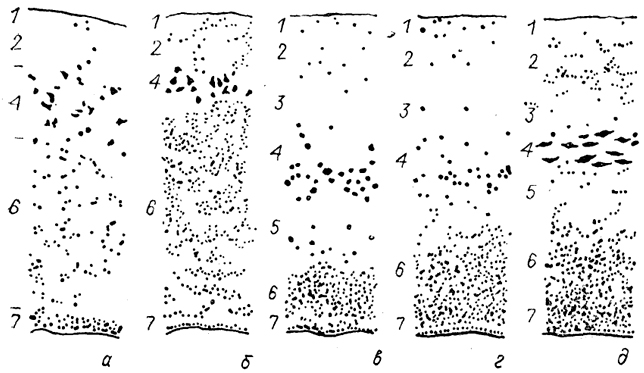
Рис. 82. Цитоархитектоника обонятельной луковицы миноги
(а), акулы
(б), двоякодышащей рыбы
(в), хвостатой амфибии
(г) и ящерицы
(д) (Nieuwenhuys., 1967).
1-7 - слои обонятельной луковицы: обонятельных волокон (1), клубочковый (2), наружный волокнистый (3), митральный (4), внутренний волокнистый (5), зернистый (6) и эпендимный (7).
У большинства позвоночных в клубочковом слое присутствуют особые тормозные нейроны - межклубочковые. Их дендриты формируют в клубочках дендро-дендритные синапсы с отростками митральных нейронов, а также контактируют с обонятельными волокнами. Аксоны этих клеток направляются в соседние клубочки, где образуют тормозные синапсы.
241
Наиболее характерным слоем обонятельной луковицы является митральный, где сосредоточены специализированные митральные нейроны. Размеры, количество, характер их распределения и соответственно степень выраженности митрального-слоя сильно варьируют у позвоночных. У одних (двоякодышащие рыбы, рептилии, птицы, млекопитающие) формируется четкий слой, у других (круглоротые, хрящевые и лучеперые рыбы, амфибии) митральные клетки расположены более или менее диффузно (рис. 82). У наземных позвоночных только один из дендритов каждого митрального нейрона входит в состав одного клубочка, тогда как у остальных - несколько дендритов принимают участие в формировании клубочков. Митральные нейроны из разных участков обонятельной луковицы связаны, по-видимому, с переработкой различной обонятельной информации. Аксоны митральных нейронов направляются в полушария.
Значительные изменения отмечены в организации зернистых, нейронов, формирующих у всех позвоночных гранулярный слой луковицы. У низших позвоночных они вместе с митральными нейронами участвуют в формировании клубочков, причем дендриты одной зернистой клетки входят в состав нескольких таких комплексов. У высших позвоночных зернистые клетки утрачивают связь как с клубочками, так и с полушарием (у них нет аксона), превращаясь таким образом в локальные интернейроны обонятельной луковицы. Дендриты зернистых клеток образуют вне зоны клубочков дендро-дендритные тормозные синапсы с митральными нейронами.
Для структуры обонятельной луковицы высших позвоночных, особенно млекопитающих, характерны особые нейроны - кисточковые. Они располагаются в наружном волокнистом слое и, подобно митральным нейронам, посылают один из дендритов в клубочек.
Эфферентные проекции обонятельной луковицы низших позвоночных сформированы аксонами митральных и зернистых нейронов, высших позвоночных - митральных и кисточковых Морфологически они оформлены в три обонятельных тракта: медиальный, латеральный и промежуточный. Последний выражен у высших позвоночных, состоит из аксонов в основном кисточковых нейронов и является комиссуральным. У других позвоночных комиссуральные связи осуществляются аксонами зернистых нейронов и проходят в составе передней комиссуры конечного мозга (у хрящевых рыб есть самостоятельный комиссуральный тракт, сходный с промежуточным обонятельным трактом высших позвоночных). Медиальный и латеральный тракты, топографически обособленные друг от друга, начинаются от разных групп митральных нейронов и проецируются в разные отделы головного мозга. У низших и высших позвоночных есть определенные различия в характере распределения окончаний обонятельных трактов (они рассматриваются в соответствующих
242
разделах главы). Здесь необходимо только подчеркнуть, что зона окончаний обонятельных проекций в структурах конечного мозга современных позвоночных ограничена.
Афферентная центрифугальная иннервация обонятельной луковицы обнаружена у всех позвоночных. В ее осуществлении принимают участие многие отделы конечного мозга (переднее обонятельное ядро, латеральный паллиум, обонятельный бугорок, ядра амигдалы), промежуточного мозга (гипоталамус, субталамус) и ряд ядер ствола (ядра шва, голубое пятно). В большинстве этих зон оканчиваются эфференты обонятельной луковицы. Центрифугальные волокна контактируют с дендритами зернистых нейронов.
Помимо основной обонятельной луковицы у ряда позвоночных (безногие и бесхвостые амфибии, многие рептилии, млекопитающие) есть дополнительная, bulbus olfactorius accessorius, которая по структуре сходна с основной и получает проекции от вомеро-назального органа (органа Якобсона). Необходимо отметить, что в дополнительной луковице даже у млекопитающих митральные нейроны располагаются диффузно, а их дендриты участвуют в формировании нескольких клубочков. Она имеет раздельные с основной луковицей зоны проекций в структурах мозга (в основном в амигдалярном комплексе), что определяет ее роль в организме.
Таким образом, обонятельные луковицы сохраняют в филогенезе позвоночных единый план строения, а их развитие идет по пути дифференциации нейронных элементов, все большего разграничения слоев и усложнения внутренних и внешних связей.
243
