Эпиталамус, Epithalamus, состоит из эпифиза (шишковидная железа, верхний мозговой придаток, Epiphysis) и поводковых (хабенулярных) ядер, nucll. habenulae.
Эпифиз в эволюции позвоночных развивался на основе так называемого теменного глаза низших позвоночных, который сформировался как производное одного или двух выростов дорсальной стенки промежуточного мозга - собственно теменного (парапинеального) органа, расположенного рострально, и пинеального органа, занимающего каудальное положение. Оба эти образования могут быть одинаково развитыми у одного и того же животного и содержать светочувствительные элементы (панцирные и костистые рыбы). У других позвоночных преобладающей структурой является пинеальный орган (круглоротые) или теменной (гаттерии, некоторые ящерицы). У высших позвоночных
236
произошла редукция теменного органа, а пинеальный, утрачивая светочувствительные элементы, превратился в железу внутренней секреции - эпифиз.
Отличительной особенностью этого отдела промежуточного мозга является его асимметрия, которую ранее пытались объяснить ранней атрофией левого парапинеального органа, однако сейчас показано, что у представителей разных групп позвоночных асимметрия комплекса разная: правосторонняя гипертрофия у круглоротых и левосторонняя - у хрящевых рыб и амфибий.
У круглоротых пинеальный и парапинеальный органы выражены различно. Пинеальный хорошо развит у миног, а у Мordacia mordax, которую считают промежуточной формой между миногами и миксинами,- отсутствует. В составе поводковых ядер выделяют перивентрикулярный и поверхностный отделы. Последний справа значительно превосходит по размерам соответствующий отдел в левых поводковых ядрах, что и определяет асимметрию комплекса. Афференты, подходящие к поводкам, идут в составе так называемых медуллярных полосок, striae medullares, и происходят главным образом из конечного мозга и частично из гипоталамуса. Эфференты формируют tr. habenulo-interpeduncularis, адресованный интерпедункулярному ядру "среднего мозга. Этот путь называют также отогнутым пучком Мейнерта, fasc. retroflexus Meynerti. В его составе имеются миелинизированные и немиелинизированные волокна.
У хрящевых рыб (например, у акулы Scyliorhinus caniculaпинеальный орган представлен длинной трубкой с дистальным закрытым отделом, лежащим на поверхности черепной коробки. Ее проксимальный конец открыт в полость III желудочка в районе между поводками и задней комиссурой. Паренхима пи-неального органа содержит фоторецепторы, опорные и ганглионарные клетки. Последние образуют пинеальный тракт, tr. pinealis, который идет к задней комиссуре и частично к поводковому комплексу. Точное место окончаний волокон не определено. Поводковые ядра составляют у хрящевых рыб ростральный отдел эпиталамуса, левые ядра комплекса превосходят правые. Характерное для большинства позвоночных разделение на медиальное и латеральное ядра у акул лишь намечается. Обе части комплекса соединяются по средней линии поводковой комиссурой, в составе которой у некоторых акул располагаются нейроны, подобные клеткам собственно ядер поводков и составляющие ядро Беллончи. У скатов оно отсутствует. Основные афференты приходят из конечного мозга (по мозговым полоскам), тектума и тегментума (текто- и тегменто-хабенулярные тракты).
У амфибий и лучеперых рыб эпиталамус состоит из комплекса дорсального и вентрального хабенулярных ядер, причем у амфибий, как отмечалось выше, наблюдается левосторонняя
237
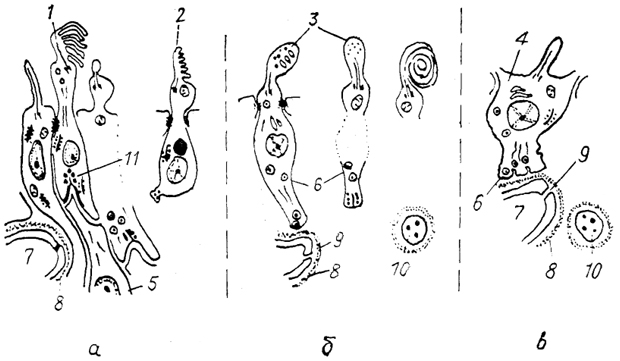
Рис. 81. Схема организации пинеалоцитов у анамний
(а), зауропсид
(б) млекопитающих
(в) (Oksche, 1988).
1 - типичный фоторецепторный пинеалоцит, 2 - серотонинсодержащий пинеалоцит, 3 - модифицированный пинеалоцит, 4 - пинеалоцит млекопитающих, 5 - нейрон, 6 - секреторные гранулы, 7 - капилляр, 8 - базальная мембрана, 9 - поры в эндотелии, 10 - симпатическое нервное окончание, 11 - специализированный синапс.
асимметрия. Эфференты поводковых ядер направляются к интерпедункулярному ядру, а часть их проходит его без переключений и адресуется глубокому и дорсальному ядрам покрышки среднего мозга.
У высших позвоночных ядра поводков выражены не столь четко, как у низших, но характер их связей остается прежним. Афференты в основном происходят из теленцефальных образований, а эфференты распределяются среди структур среднего мозга: в интерпедункулярном ядре, ретикулярной формации, а также (у млекопитающих - от латерального ядра) в области мезенцефалического ядра шва.
У высших позвоночных прослеживаются регрессивные изменения в структуре эпифиза. Они выявляются в пределах каждого класса и особенно среди млекопитающих. Их эпифиз прикрепляется к поводковому комплексу и содержит клетки, получившие наименование пинеалоциты, а также интерстициальные клетки, подобные глиальным. Полагают, что пинеалоциты являются дериватами сенсорных клеток (рис. 81), которые опосредовали воздействие световых раздражителей на эндокринную систему, и таким образом являлись фотонейро-эндокринными элементами.
Смена функций, приведшая к превращению пинеального органа в железистое образование - эпифиз, определила значительные изменения и в структуре основных клеток. У птиц и млекопитающих
238
наиболее значительные изменения произошли в наружных сегментах пинеалоцитов. В результате у птиц и млекопитающих эти клетки, как полагали, утратили способность к реакции на свет. Но в последнее время показано, что пинеалоциты птиц сохранили способность к синтезу специфических фоторецепторных белков и реакции на свет. Пинеалоциты птиц и млекопитающих - типичные секреторные железистые элементы, синтезирующие мелатонин. Окончательным доказательством единства происхождения фоторецепторных пинеалоцитов низших позвоночных и секреторных клеток эпифиза высших позвоночных послужили иммуногистохимические данные о присутствии в типичных фоторецепторных пинелоцитах костистых рыб серотонина - предшественника мелатонина - гормона эпифиза.
Пока неясен механизм влияния секреторных клеток эпифиза птиц и млекопитающих на циркадную ритмику физиологических процессов, поскольку они непосредственно не подвергаются: воздействию световых раздражителей, но, возможно, это опосредуется через взаимодействие с ядрами гипоталамуса, получающими ретинальные входы (например, супрахиазматическое ядро). Нарушение этих связей приводит к нарушению циркадной; ритмики в работе эндокринных систем, связанных с эпифизом.
В целом структуры эпиталамуса в достаточно дифференцированном виде представлены уже у самых примитивных из ныне живущих позвоночных и на протяжении эволюции характеризуются постоянством состава и выполняемых функций. Так, пинеальный и парапинеальный органы, содержащие у низших, экстраретинальные фоторецепторы, обеспечивают контроль циркадной ритмики многих физиологических процессов. Эпифиз млекопитающих и птиц, лишенный рецепторных клеток, за счет гипоталамических входов продолжает выполнять роль, свойственную этому отделу мозга. Ядра поводков у всех позвоночных животных дают начало хабенуло-интерпедункулярной системе волокон. Последняя рассматривается как экстрагипоталамическая система пищевого поведения, а у высших как существенное, звено лимбической системы.
239
