Гипоталамус, Hypothalamus, занимает самое вентральное положение среди диенцефальных образований. Он представлен нервной тканью, окружающей нижнюю часть III желудочка и сосредоточенной в основании мозга. Относительно малые размеры этого отдела, отсутствие крупных миелинизированных трактов, диффузное распределение нейронов, составляющих непрерывную клеточную массу с различной степенью концентрации элементов, - все эти черты характерны для гипоталамуса позвоночных независимо от их таксономической принадлежности и обусловливают объективные сложности классификации гипоталамических образований даже в пределах одной группы.
Наиболее часто структуры гипоталамуса разделяют (даже у млекопитающих) на зоны в зависимости от их удаленности от вентрикулярной стенки: перивентрикулярную, медиальную и латеральную. Степень развития разных отделов гипоталамуса варьирует: у многих низших позвоночных основные клеточные элементы сосредоточены в перивентрикулярной области, у высших представителей отдельных таксономических групп пластиножаберные,
230
костистые рыбы, рептилии) дифференциация структур гипоталамуса идет прежде всего за счет медиальной зоны, а у птиц и млекопитающих - за счет латеральной.
В составе гипоталамуса часто выделяют в качестве самостоятельного отдела преоптшескую область (АР), занимающую наиболее ростральное положение в этом отделе мозга (рис. 67). Ее часто рассматривают как неэвагинировавшую часть конечно-то мозга. Особенно хорошо данная область выражена у низших позвоночных. У низших млекопитающих в ней выделяют те же зоны, что и в остальной части гипоталамуса: латеральную, медиальную и перивентрикулярную. У приматов данная область дифференцирована крайне слабо.
У низших позвоночных гипоталамус образован тонкостенной структурой, лежащей в основании мозга, конфигурация которой зависит от формы III желудочка - многочисленные углубления вентрикулярной полости приводят к изменению очертаний этого отдела и вместе с перекрестом зрительного нерва часто служат ориентирами для разделения гипоталамуса на области или отделы (рис. 65, 67).

Рис. 79. Основные связи нижней доли гипоталамуса ската
Raja eglanteria (Smeets, Boord, 1985).
1 - паллиальный тракт, 2 - базальная поверхностная область, 3 - МПП, 4 - лобо-церебеллярный тракт. X - перекрест волокон.
У многих круглоротых основные клеточные группы сосредоточены вблизи желудочка. У миноговых, хрящевых и лучеперых рыб в составе гипоталамуса появляются образования, удаленные от вентрикулярной стенки. Специфическим отделом мозга этих животных является крупная нижняя доля гипоталамуса, lobus inferior hypothalami.
Нижняя доля - структура гетерогенная, включающая несколько клеточных групп (рис. 66). Данные относительно ее связей фрагментарны. Показано, что у рыб с высокодифференцированной вкусовой системой этот отдел мозга получает входы от вторичного вкусового ядра или от его гомолога-ядра F (у пластиножаберных рыб). Другие афференты поступают от тегментальной области и тектума среднего мозга, мозжечка, а у пластиножаберных рыб - от контрлатерального полушария конечного мозга (рис. 79). Эфференты нижней доли у пластиножаберных
231
и костистых рыб направляются к моторным структурам ствола мозга. В последнее время подтверждено существование у пластиножаберных рыб связей нижней доли с мозжечком - это лобо-церебеллярный тракт, описанный ранее. Он начинается в каудальной части нижней доли и адресуется передней части тела мозжечка. Физиологические данные свидетельствуют об участии нижней доли в регуляции пищевого поведения у скатов.
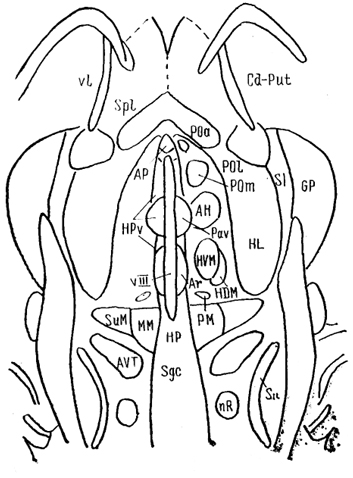
Рис. 80. Ядра гипоталамуса мозга крысы на горизонтальном срезе (Swanson, 1987).
АН - переднее гипоталамическое, Аг - аркуатное, РМ - премамиллярное, ММ - мамиллярное, SuM - супрамамиллярное, РОа - переднее, Pol - латеральное и Рот - медиальное преоптические ядра, SI - безымянная субстанция.
Остальные отделы гипоталамуса рыб во многих отношениях сходны с таковыми наземных позвоночных различия заключаются лишь в степени развития каждой из зон.
У всех позвоночных перивентрикулярная зона гипоталамуса (HPv) организована довольно просто в состоит из примитивных клеток изодендритного типа. Нейроны образуют узкую клеточную полоску примыкающую к стенке желудочка и пронизанную многочиеленными волокнами перивентрикулярной системы. Наиболее заметной клеточной агрегацией этой зоны является паравентрикулярное ядро (nucl. paraventricularis), которое сохраняет свое положение и в мозге других позвоночных (рис. 69, 80).
Общая черта всех позвоночных - это наличие в данной зоне нейросекреторных клеток, а также нервных элементов, получающих прямой вход от волокон зрительного нерва. Сама зона тесно связана со структурами циркумвентрикулярной системы в области III желудочка (в частности, у низших позвоночных здесь широко представлены клетки-таннициты, расположенные в срединном возвышении и способные к активному транспорту и абсорбции нейрогормонов). Число нейросекреторных клеток особенно велико у низших позвоночных в преоптической перивентрикулярной области, а у наземных - в супраоптическом и: паравентрикулярном ядрах. Их аксоны образуют гипоталамо-гипофизарный тракт и выделяют нейрогормоны и рилизинг-факторы
232
в отделы гипофиза. Тем самым обеспечивается; прямая связь мозга с эндокринной системой.
Другой чертой, характерной для перивентрикулярной зоны гипоталамуса, является наличие клеток-мишеней зрительных афферентов. Зрительные проекции адресованы, и передней части) медиальной зоны. Они описаны у всех исследованных видов, позвоночных, но районы их окончания обозначаются по-разному. У хрящевых рыб структура, получающая такие входы, обозначалась разными авторами как гипоталамическое зрительное ядро, хиазматическое ядро, супрахиазматическое ядро.* Под последним, наиболее распространенным, названием оно описано у многих представителей позвоночных. Полагают, что у всех позвоночных эта структура гипоталамуса связана с регуляцией циркадной ритмики различных процессов в. организме, в том числе циклов сон - бодрствование - у высших. У млекопитающих помимо входа от сетчатки сюда адресованы проекции ЛКТ, а эфференты связывают это ядро с эпифизом, оказывая влияние на продуцирование последним гормонов.
Таким образом, перивентрикулярная зона гипоталамуса имеет сходное строение у всех позвоночных, в основном содержит нейроны, обеспечивающие связи гипоталамуса с эндрокринными органами и обусловливает регуляцию циклических процессов, зависящих от степени освещенности.
У наземных позвоночных наиболее обширна медиальная зона гипоталамуса (НМ). Она состоит из нескольких ядер, частей разделяемых в соответствии с расположением в рострокаудальном направлении на ядра передней, средней и задней групп. Ядра передней группы (SO, Sch) содержат нейросекреторные клетки и нейроны, получающие зрительные афференты из сетчатки, и оказываются в структурном и функциональном отношениях сходными с элементами перивентрикулярной зоны. В средней группе основными являются вентро- и дорсомедиальное ядра гипоталамуса (HVM, HDM). Задняя группа (HP) слабо выражена у низших позвоночных и только намечается у хвостатых амфибий. Вообще у анамний и рептилий супраоптическое и супрахиазматическое ядра имеют диффузное строение, а зона НМ слабо намечена. При этом некоторые ядра (например, супраоптическое) присутствуют почти у всех позвоночных, другие (например, некоторые ядра задней группы) дифференцируются только у млекопитающих.
Основной вход в ядра медиальной зоны (за исключением передней группы) составляют отделы лимбической системы конечного мозга (амигдала, септальная область), а также сенсорные ядра ствола мозга, обеспечивающие проведение информации от вкусовых, висцеральных, обонятельных, зрительных, а
233
у млекопитающих и от соматических и слуховых рецепторов. Эфференты адресованы соседним, перивентрикулярной и латеральной, зонам гипоталамуса, а также - в составе медиального пучка переднего мозга - лимбическим структурам, ретикулярной формации, району центрального серого вещества среднего мозга и, наконец, премоторным и вегетативным ядрам ствола.
В функциональном отношении охарактеризовать структуры медиальной зоны довольно сложно. Очевидно, что их совокупная деятельность обеспечивает участие этой области промежуточного мозга в организации сложных форм пищевого, питьевого, полового и т. п. поведения, что обусловлено как наличием разнообразных сенсорных входов, так и множественными выходами на исполнительные системы организма (через перивентрикулярную зону - на эндокринную систему, через премоторные и вегетативные ядра - на сомато- и висцеромоторные образования) и к лимбическим структурам конечного и среднего мозга.
Среди всех вышеназванных структур медиальной зоны выделяются мамиллярные тела, corpora mammillares, которые хорошо выражены у высших позвоночных. У млекопитающих они представлены крупными выпуклыми образованиями, лежащими на вентральной поверхности мозга и состоящими из нескольких ядер. Система их связей оказывается иной, чем у других ядер медиальной зоны. Основной вход к мамиллярным телам образован гиппокампальными нейронами, аксоны которых у млекопитающих образуют крупный проводящий тракт - свод, fornix. Сенсорные афференты, в отличие от других ядер, приходят сюда из сенсорных полей (зрительной и слуховой областей) новой коры. Характер распределения эфферентов также различается: основные проекции адресованы ядрам передней группы таламуса, а через них - лимбической области коры. Система внутриги-поталамических связей у мамиллярных тел не развита. Полагают, что мамиллярные тела и связанные с ними образования имеют отношение к организации процессов обучения и памяти.
Необходимо отметить, что у млекопитающих в составе мамиллярного комплекса часто описывают группу клеток, которая в ходе эволюции этого класса позвоночных вычленяется в самостоятельный крупный комплекс - ядра серого бугра, или туберальные ядра. Слабо выраженный у низших млекопитающих, он значительно эволюционирует среди приматов, а у антропоидов и человека имеет в своем составе ядра, отсутствующие у других приматов. Учитывая тесные связи данных ядер с корой, полагают, что развитие этого отдела шло параллельно с развитием некоторых отделов новой коры.
Нейронные группы, лежащие среди волокон медиального пучка переднего мозга, составляют латеральную зону гипоталамуса (HL). У птиц и млекопитающих она достигает значительных размеров и дифференцируется на ряд полей и ядер. У антропоидов в ее пределах дополнительно выделяется латеральная
234
преоптическая зона, латеральное и супрамамиллярное ядра, отсутствующие у других высших позвоночных. В силу особенностей строения и связей эту зону часто рассматривают как .диенцефальный компонент ретикулярной формации. Об этом же свидетельствуют результаты физиологических наблюдений, в частности стимуляция этой области приводит к реакции активации, характерной для восходящей ретикулярной формации. Относительно афферентов этой области сведения немногочисленны. Эфференты распределяются почти по всей поверхности новой коры, в лимбических структурах конечного мозга, а также в моторных и вегетативных ядрах ствола мозга.
Основные связи гипоталамуса сходны у всех позвоночных. Большинство афферентов и эфферентов этой области проходит в составе многокомпонентного медиального пучка переднего мозга (МППМ, fasciculus prosencephali medialis). В мозге млекопитающих он связывает более чем 50 образований, начиная от ростральных отделов конечного мозга и кончая спинным. Значительное число эфферентов входит в состав перивентрикулярной системы волокон, связывающих гипоталамус с таламусом, средним мозгом, перивентрикулярно расположенными отделами ствола. Собственной системой связей гипоталамуса является гипоталамо-гипофизарный тракт, реализующий воздействие на эндокринную систему. Связи со структурами старой коры образованы у наземных позвоночных специфической системой волокон - сводом.
Таким образом, во многих отношениях гипоталамус оказывается консервативным отделом мозга, сохраняющим сходные черты организации у всех позвоночных. Наиболее ярко это проявляется в строении и системе связей перивентрикулярной зоны. Вместе с тем в гипоталамусе возникают черты прогрессивной дифференцировки, которая выражается у высших рыб в развитии нижней доли, а у птиц и млекопитающих - латеральной зоны. В то же время у других представителей позвоночных (низшие лучеперые рыбы, двоякодышащие, амфибии) сохраняется исходное, перивентрикулярное положение основных элементов гипоталамуса.
Несмотря на многочисленные исследования различия в подходах не позволяют провести гомологизацию многих отделов гипоталамуса разных позвоночных. Однако в тех случаях, когда это возможно, четко прослеживаются черты прогрессивных преобразований в ряде отделов гипоталамуса у млекопитающих по сравнению с птицами, а среди млекопитающих - у приматов по сравнению с другими группами. Это выражается прежде всего в появлении у приматов в пределах задней группы ядер медиальной зоны дополнительных объединений и в дифференциации латеральной зоны (при сравнении с птицами). На нейронном уровне прогрессивные тенденции в развитии гипоталамуса сказываются в появлении у некоторых млекопитающих
235
тенденции к сосредоточению нейронов в пространственные комплексы-кластеры.
В целом же в составе гипоталамуса выделяют аналогичные области, элементы которых, имея сходные связи, вероятно, выполняют и сходные функции у разных позвоночных. Это, во-первых, обилие выходов на эндокринную систему, реализуемых через гипофиз и в основном осуществляемых элементами перивентрикулярной области. Во-вторых, конвергенция разномодальной импульсации (у низших, в основном, висцеросоматической, а у высших и экстероцептивной) и множество доступов к исполнительным отделам мозга (моторным и вегетативным ядрам), обеспечивающие участие гипоталамуса в организации поведенческих реакций, в том числе сложных комплексных форм поведения (у скатов, птиц и млекопитающих). В-третьих, наличие у всех позвоночных ретинальных входов в гипоталамус, что, вероятно, представляет собой реализацию древнего механизма регуляции жизнедеятельности организма в зависимости от степени освещенности и времени суток и осуществляется за счет влияния гипоталамуса на гипофиз, а у высших позвоночных и на эпифиз.
Наконец, согласно физиологическим данным, в гипоталамусе млекопитающих обнаружены так называемые "центральные" рецепторы, активация которых возможна не синаптическим путем, а сенсорными стимулами (центральные глюко- и терморецепторы). В ходе эволюции происходит дифференциация структур гипоталамуса, приводящая к формированию участков, связанных с высшими отделами мозга и вовлеченных таким образом в регуляцию и обеспечение сложных форм поведенческих реакций. Отметим, что у млекопитающих гипоталамус рассматривается в качестве "выходного" звена лимбической системы именно с учетом характера его связей.
236
*
У хрящевых рыб Platyrhinoidis зрительный вход- обнаружен в каудальном гипоталамусе на уровне заднего бугорка.
