Область покрышки среднего мозга и перешейка содержит значительное число образований, далеко не для всех из которых установлены возможные гомологи и выяснены пути преобразования в ходе эволюции. Одна из причин этого - ретикулярное происхождение большинства этих структур: одни тегментальные ядра включены в состав ретикулярной формации, другие, хорошо развитые в мозге высших позвоночных, вероятно, ее производные. Таким образом, становятся понятными многочисленные разночтения и противоречия, имеющиеся в литературе, которая посвящена описанию этих отделов у разных позвоночных.
Тем не менее в составе тегментума перешейка и среднего мозга можно выделить несколько групп образований, сходных в отношении связей и выполняемых функций. Одну из таких групп составляют структуры, обеспечивающие проведение импульсации к моторным ядрам черепномозговых нервов и к спинному мозгу - так называемые премоторные структуры, или надсегментарные моторные отделы. Несмотря на то, что у низших позвоночных эти отделы принадлежат ретикулярной формации, они в отличие от других ретикулярных образований четко представлены в мозге разных позвоночных и могут быть гомологизированы.
В пределах базальной пластинки среднего мозга высших позвоночных располагается крупное скопление клеток - красное ядро, nucl. ruber (рис. 59). Полагают, что данная структура свойственна всем тетраподам, и ее специфической характеристикой являются клетки - источники рубро-спинального тракта, tr. rubrospinalis. Для высших позвоночных идентификация этой структуры не вызывает затруднений. У других же при отсутствии видимых клеточных скоплений (при обработке по Нис-слю), тем не менее в области тегментума можно выделить группы клеток со сходными связями. Поэтому в настоящее время для выделения примордиального красного ядра используют несколько критериев: положение в мозге, наличие входов в составе верхних ножек мозжечка и контрлатеральный выход на спинной мозг.
Перекрещенный рубро-спинальный тракт описан у бесхвостых амфибий, четвероногих рептилий, птиц и млекопитающих. Считалось, что у примитивных животных подобные связи отсутствуют. Действительно, у круглоротых не обнаружено тегменто-
168
спинальных связей, аналогичных рассматриваемым, однако у некоторых рыб в пределах тегментума выделяется группа клеток - источников спинальных связей. Кроме того, у некоторых видов скатов (Raja clavata, Dasyatus sabina) не только четко выделяется группа клеток - примордиум красного ядра, но развиты входы от ядер мозжечка и хорошо выражен перекрещенный рубро-спинальный тракт (заметим, что речь идет о рыбах, интенсивно использующих для движения плавники), о б в
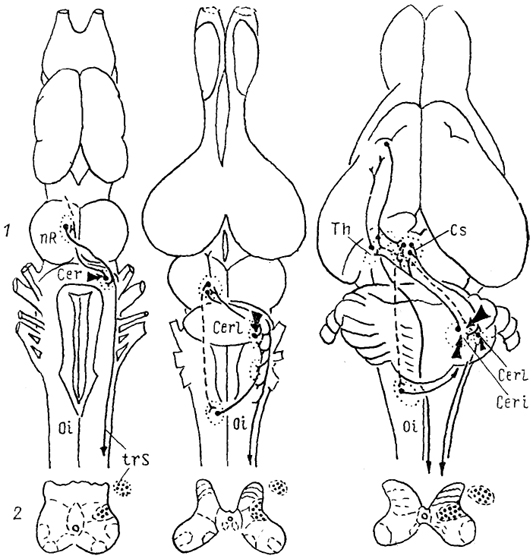
Рис. 59. Эволюция красного ядра (Toth e. а., 1985, Donkelaar, 1988).
а-в - красное ядро бесхвостой амфибии (а), ящерицы (б), опоссума (в). 1 - связи красного ядра, 2 - окончания волокон рубро-спинального тракта.
В мозге бесхвостых амфибий красное ядро выделяется на основе анализа связей: группа клеток, лежащих в вентро-медиальной части тегментума, получает входы от единственного ядра мозжечка через слабо развитые верхние ножки мозжечка и: формирует перекрещенный рубро-спинальный тракт. Пожалуй, бесхвостые амфибии - самые примитивные из групп, обладающих типичным красным ядром. При этом степень развития моторных систем мозга у них далеко не совершенна. Связи с нижней оливой отсутствуют.
У рептилий красное ядро варьирует по размерам. У некоторых
169
групп оно не может быть цитоархитектонически выделено, у других же хорошо развито, например у четвероногих, и в его составе выделяются клеточные группы, четко отграниченные от окружающей ретикулярной формации. Мозжечковые входы к нему образует латеральное ядро мозжечка. Кроме того, формируются связи от вышележащих отделов - ядер конечного мозга. Эфференты сосредоточены в крупных рубро-бульбарном и рубро-спинальном трактах. Последний проходит через дорсальную часть латерального канатика и оканчивается в латеральном отделе пластин V-VI. В случае отсутствия рубро-спинального сохраняется рубро-бульбарный тракт (например, у питона). Таким образом, у рептилий, особенно имеющих конечности, наблюдается дальнейшее развитие системы красного ядра. Однако связи с нижней оливой на этом уровне еще слабо представлены, равно как и входы от вышележащих уровней мозга.
У птиц в составе красного ядра выделяется по крайней мере два типа нейронов: крупные, сосредоточенные преимущественно дорсомедиально и вентролатерально, и средние (и мелкие), сосредоточенные в основном рострально. Характер связей подобен имеющимся у рептилий.
Только у млекопитающих в пределах данного ядра клетки разных размеров распределены в различных областях структуры, в результате чего в пределах красного ядра описывают крупноклеточную и мелкоклеточную части, partes magno-et parvocellularis. Удельный вес последней и ее связей прогрессивно возрастает, при этом крупноклеточная часть у высших приматов и человека в значительной степени редуцируется. У сумчатых же основной объем красного ядра составлен крупными клетками, а мелкие нейроны еще не сосредоточены в самостоятельную группу. Степень развития красного ядра коррелирует и со способом локомоции: способность к плаванию и полету сочетается с относительно незначительно развитой структурой, а хождение и длинные конечности - с большим по размерам красным ядром и обширными связями его крупноклеточной части. Входы к красному ядру млекопитающих берут начало от разных ядер мозжечка. Уже в мозге примитивных млекопитающих афференты промежуточного, и латерального (зубчатого) ядер мозжечка адресованы разным отделам. Появляются входы от коры мозга. Эфференты также происходят из разных областей: так, у опоссума рубро-спинальный тракт образован крупными клетками, лежащими в каудальной и ростровентральной областях; ростромедиальная и ростродорсальная же образуют рубро-оливарные и рубро-бульбарные проекции соответственно.
У приматов и человека ростральная, наиболее значительная часть, состоящая из относительно мелких клеток, получает входы от новой коры, стриатума и зубчатого ядра мозжечка, а
170
проецируется на главное ядро нижней оливы и особую группу таламйческих ядер. Крупноклеточная часть имеет незначительное число входов от конечного мозга, основной мозжечковый вход образован нейронами промежуточного ядра. Ее эфференты составляют очень небольшой рубро-спинальный тракт, распределение которого ограничивается верхними сегментами спинного мозга. Очевидно, в ряду млекопитающих происходит постепенное возрастание роли мелкоклеточной части красного ядра, что, вероятно, связано с общим усложнением моторных систем мозга и развитием высших моторных систем, берущих начало с уровня конечного мозга.
Таким образом, крупноклеточная часть красного ядра млекопитающих и в целом красное ядро других позвоночных представляют собой надсегментарный уровень организации моторных систем. Вместе с вестибуло- и ретикуло-спинальным трактами и медиальным продольным . пучком рубро-спинальный тракт обеспечивает организацию многих моторных реакций, в частности локомоции. Развитие же мелкоклеточной части красного ядра, происходящее в связи с усложнением вышележащих отделов, является следствием развития в эволюции высших моторных систем (пирамидной и экстрапирамидной), объединяющих многие отделы мозга.
Отметим, что в мозге круглоротых и всех групп рыб достаточно четко выделяется группа нейронов, образующая моторное ядро покрышки, nucl. motorius tegmenti. Оно является местом конвергенции входов от разных структур, в том числе ют тектума и торуса. Его эфференты проецируются на моторные ядра ствола и спинного мозга. Очевидно, это свойственная низшим премоторная структура аналогична красному ядру; некоторые же авторы рассматривают ее как прекурсор последнего.
Другими структурами, которые можно отнести к премоторным образованиям являются ядро медиального продольного пучка, nucl. fasc. longitudinalis medialis, и промежуточное ядро Кахаля, nucl. interstitialis Cajal. На их нейронах конвергируют входы от разных источников, в том числе от вестибулярного комплекса, тектума (передних холмов), претектальной области. Эфференты формируют часть медиального продольного пучка (МПП), а у некоторых животных - направляются и к ядрам глазодвигательного комплекса. МПП - многокомпонентный путь, свойственный мозгу всех позвоночных. Он соединяет ствол мозга с моторными отделами спинального уровня, характеризуется постоянством расположения и занимает парамедианную позицию в стволе, проходя в вентральных канатиках спинного мозга на большое расстояние. Промежуточное ядро и ядро Кахаля - не единственные, образующие данный путь; на уровне среднего мозга его составляют также эфференты ядра задней комиссуры, а у высших позвоночных и
171
ядра Даркшевича, и красного ядра. На уровне ромбовидного мозга к нему присоединяются вестибуло-спинальные волокна, а также незначительное число волокон, источниками которых являются сенсорные ядра черепномозговых нервов и мозжечка (показано у рептилий).* Множественность связей определяет участие МПП в таких, реакциях, как сочетанный поворот головы и глаз.
На уровне среднего мозга у разных групп позвоночных имеются и другие структуры, связанные с моторными отделами, однако недостаточность сведений о связях и функциональной роли этих структур не позволяет судить об их гомологичности с топографически сходными ядрами других животных.
Так, у многих позвоночных описано глубокое ядро среднего мозга, nucl. profundus mesencephali, достигающее у некоторых групп значительных размеров (например, у рептилий). Согласно имеющимся данным у акул, амфибий и: рептилий оно получает билатеральный тектальный вход и проецируется на нижележащие моторные структуры. Относительно других позвоночных данные достаточно противоречивы.
В тегментальной области перешейка пластиножаберных описано несколько клеточных групп, обозначенных как ядра F, G, Н. Два первых проецируются на спинной мозг, последнее, возможно, опосредует текто-бульбарные связи, однако недостаточность сведений не дает возможности говорить об их специфичности или гомологии с какими-либо центрами других животных.
Возможно, к этой же группе относится ядро, обнаруженное у всех актиноптеригий и расположенное в вентролатеральной части покрышки. У представителей разных групп лучеперых оно описано под разными названиями: красное ядро покрышки, nucl. ruber tegmenti, латеральное ядро торуса, дорсальная часть энтопедункулярного ядра, nucl. entopeduncularis pars dorsalis. Степень развития ядра варьирует, связи изучены недостаточно, хотя имеются сведения о наличии его эфферентов в составе лобо-бульбарного и лобо-церебеллярного трактов. Отметим, однако, что анализ его связей у голостей позволил предположить, что данная структура является частью торуса.
В числе структур, обеспечивающих связи стволовых отделов; с моторными центрами, у наземных позвоночных выделяются образования, связанные с вокализацией. К ним относится описанное у бесхвостых амфибий nucl. pretrigeminalis (у Хепоpus - дорсальная тегментальная область). Оно располагается в латеральной части тегментума перешейка. Для его нейронов-характерны высокая зависимость от уровня гормонов, выраженные связи с преоптической областью гипоталамуса и наличие
172
эфферентов, адресованных билатерально ядрам языко-глоточного и блуждающего нервов.
У птиц вентромедиально от желудочка среднего мозга располагается дорсомедиальное интерколликулярное ядро, которое получает афференты от слухового мезенцефалического отдела и адресует эфференты каудальной части ядра подъязычного нерва, осуществляющей иннервацию голосового аппарата.
Помимо премоторных образований на уровне среднего мозга располагаются структуры, выполняющие релейную роль и связывающие между собой различные центры. Такую роль приписывают, в частности, ядру задней комиссуры, nucl. commissurae posterioris, которая у высших позвоночных тесно связана с тектумом. У птиц его вероятным гомологом является ядро, .лежащее в претектальной области, - nucl. spiriformis lateralis thalami. Значительная часть входов к этой структуре у птиц и рептилий образована стриатумом. Таким образом, учитывая особенности контактов, этот отдел рассматривают как звено, опосредующее стрио-тектальные связи, т. е. передающее влияние высших отделов на премоторное образование - глубокие слои тектума, дающие начало нисходящим проекциям. У млекопитающих ядро задней комиссуры обеспечивает связи в пределах ядер глазодвигательного нерва и, как полагают, опосредует движения глаз вверх. Афференты от стриатума не обнаружены, и считается, что аналогичную роль у млекопитающих играет черная субстанция.
Черная субстанция, substantia nigra, - характерная для мозга млекопитающих структура, лежащая в вентральной части покрышки. Она разделяется на компактную и ретикулярную части, partes compacta et reticulata, соответственно. Компактная часть у человека и высших приматов имеет специфическую пигментацию, обусловленную наличием дофаминергических нейронов (у других млекопитающих менее выражена). Тем не менее локализация ядра у всех млекопитающих сходна: вентральная часть среднего мозга у низших млекопитающих, не имеющих компактных ножек мозга, и положение между тегментумом и ножками мозга у видов, где последние хорошо развиты. Каудально черная субстанция простирается до мостовых ядер. Связи двух отделов черной субстанции различны. Ретикулярная часть получает входы от стриатума и проецируется в тектум, таламус и педункуло-понтийное ядро. Компактная часть имеет множество источников афферентации, среди которых миндалевидный комплекс, ядра шва, хабенулярные ядра. Эфференты адресованы стриатуму. Таким образом, именно ретикулярная часть черной субстанции может рассматриваться в качестве аналога вышеупомянутых ядер, опосредующих стрио-тектальные связи. В целом же черная субстанция расценивается как одно из важнейших звеньев экстрапирамидной системы млекопитающих.
173
Гомологом черной субстанции у птиц, рептилий и бесхвостых амфибий считают покрышечное педункуло-понтийное ядро, nucl. tegmenti pedunculopontinus, лежащее в вентролатеральной области среднемозгового тегментума; у некоторых рептилий (например, ящерицы) его называют, как и у млекопитающих, черной субстанцией. Связи этого ядра характеризуются двусторонними проекциями к стриатуму и таламусу, а также зфферентами, адресованными тектуму и ретикулярной формации ствола. Помимо сходства связей имеется и сходство-нейрохимических характеристик: в составе данного ядра обнаружены катехоламинергические нейроны. При этом у птиц, они сосредоточены в компактной части педункуло-понтийного' ядра, которую считают гомологом одноименного отдела черной; субстанции, гомологом же ретикулярной части у них является дорсальное покрышечное педункуло-понтийное ядро.
Сведения относительно гомологичных черной субстанции структур в мозге других позвоночных фрагментарны. Так, у акул в ростральной части среднего мозга находится скопление-мелких нейронов - латеральное ядро покрышки, nucl. tegmenti lateralis.* Известно, что оно имеет двусторонние связи с тектумом и афференты от спинного мозга. Некоторые авторы рассматривают латеральное ядро покрышки как примордиум черной субстанции.
Покрышечное педункуло-понтийное ядро описано и в мозге млекопитающих, однако по характеру связей оно отличается от одноименного ядра других позвоночных тем, что адресованные ему афференты происходят из вентральной части стриатума и ретикулярной части черной субстанции. Кроме того, характерными являются двусторонние связи с субталамическим ядром. Не вызывает сомнений принадлежность покрышечного педункуло-понтийного ядра к экстрапирамидной системе млекопитающих. Некоторые авторы в качестве предшественника этого отдела рассматривают интерколликулярное ядро рептилий, которое характеризуется входами от вентральной части стриатума и эфферентами к ретикулярной формации и тектуму.
Таким образом, часть структур среднего мозга, расположенных в области покрышки, по-видимому, является звеньями многокомпонентных систем мозга, в частности моторных. У млекопитающих эти структуры наиболее подробно изучены, о других животных сведения в основном фрагментарны. Недостаточность представлений о системной организации затрудняет гомологизацию входящих в эти структуры образований. Отметим, впрочем, что эта организация может значительно отличаться: если у млекопитающих важнейшим премоторным центром, передающим влияния теленцефальных отделов на спинной мозг, является крупноклеточная часть красного ядра,
174
то у других, особенно у высших позвоночных, эта роль может принадлежать глубоким слоям тектума, и таким образом сами, системы окажутся состоящими из разных компонентов - ситуация, в которой, вероятно, установление гомологии нельзя считать правомочным.
На уровне среднего мозга имеется структура, свойственная мозгу всех ныне живущих позвоночных, гомологичность которой не вызывает сомнений, хотя наши представления относительно ее роли далеко не полны. Речь идет об межножковом, или интерпедункулярном ядре, nucl. interpeduncularis. Оно хорошо выражено уже в мозге круглоротых и хрящевых рыб, где располагается в области шва на границе перехода перешейка в средний мозг. Описания этого ядра у пластиножаберных противоречивы, поскольку не все авторы его разграничивают от ядер шва. У высших позвоночных интерпедункулярное ядро латерально ограничено ножками мозга. У всех позвоночных ядро образовано мелкими нейронами с небольшим объемом перинуклеарной цитоплазмы. В пределах структуры размеры нейронов варьируют, что является одним из оснований для выделения нескольких клеточных групп: так, у кошек выделяют 4 или 5, а у крыс от 4 до 7. У миног и акул деление структуры производят по топографии: ромбенцефалическая и мезенцефалическая части у миног, дорсальная и вентральная - у акул. Медиаторные характеристики нейронов ядра также служат основанием для разделения его структуры.
В мозге всех позвоночных основной вход к интерпедункулярному ядру образуют нейроны поводковых (хабенулярных) ядер эпиталамуса. Их аксоны формируют центральную часть отогнутого пучка Мейнерта, fasc. retroflexus Meynert, и, сложным образом перекрещиваясь, входят в межножковое ядро. В исследованных случаях обнаружен сложный и упорядоченный характер организации хабенуло-интерпедункулярных связей. Так, в интерпедункулярном ядре бесхвостых амфибий афференты пронизывают определенным образом ориентированные дендриты нейронов, что приводит к активации практически всей структуры в ответ на стимуляцию одной части хабенуло-интерпедункулярного тракта. Четкая топологическая закономерность обнаружена и в организации связей медиального ядра поводков и интерпедункулярного ядра у млекопитающих.
Другие источники входов в интерпедункулярное ядро известны далеко не в полной мере. Для некоторых животных обнаружены входы от тектума. У млекопитающих среди источников афферентации ядро диагональной полоски Брока, дорсальное ядро покрышки, мезенцефалическое ядро шва, центральное серое вещество и голубое пятно.
Эфференты интерпедункулярного ядра у двоякодышащих рыб и хвостатых амфибий образуют интерпедункуло-бульбарный тракт, отсутствующий у лучеперых рыб. У анура эфференты
175
ядра направляются к дорсальному и глубокому ядрам покрышки. У млекопитающих, изученных в этом отношении более детально, они адресованы дорсальному ядру покрышки, медиодорсальному ядру таламуса, септальным ядрам, переднему ядру мамиллярных тел, ядру диагональной полоски Брока, преоптической области, вентральному ядру покрышки и латеральному ядру поводков. У некоторых видов обнаружены лроекции в гипоталамус и к центральному ядру шва.
Характерной особенностью нейронов данной структуры является значительное число нейросекреторных элементов. Они обнаружены у некоторых видов, относящихся к разным классам, и, возможно, эта черта свойственна всем позвоночным. Так, у лягушек нейросекреторные клетки содержат крупные гранулы, а отростки имеют крупные варикозные расширения. Дендриты, ветвящиеся в плоскости, перпендикулярной пучку Мейнерта, образуют по всей длине щелевые контакты и заканчиваются на субпиальной поверхности, контактируя с межножковой цистерной. Подобные клетки описаны и в мозге человека, где их отростки формируют концевые ножки на сосудах и пиальной оболочке. Наличие нейросекреторных элементов, а также сильная васкуляризация ядра (по-видимому, свойственная всем позвоночным, имеющим систему кровоснабжения мозга), позволяют объяснить системный характер влияний этой структуры на различные формы поведенческих реакций. Действительно, сначала только на основании анализа связей, а позднее - экспериментальных исследований, был сделан вывод о принадлежности межножкового ядра к структурам лимбической системы, которые опосредуют передачу импульсации на премоторные центры - дорсальное и глубокое тегментальные ядра. Показано участие этих отделов в обеспечении избегания, полового и (у высших) эмоционального поведения. Раннее созревание этого ядра в онтогенезе позволило предположить, что ряд поведенческих реакций новорожденных млекопитающих (например, сосание и глотание) также опосредуется активацией этого ядра. Полагают, что одним из способов осуществления влияний интерпедункулярного ядра на другие структуры мозга может быть выброс биологически активных веществ (в частности, соматостатина) в церебро-спинальную жидкость. Отметим, что довольно сложной оказывается и нейрохимическая характеристика интерпедункулярного ядра; здесь обнаружены нейроны и терминали различной медиаторной природы. Высказывается предположение, что в ряду позвоночных, особенно млекопитающих, происходит нейрохимическая специализация ядра, что может отражать общее прогрессивное развитие лимбической системы у высших позвоночных. Однако недостаточность сведений не позволяет окончательно сделать такой вывод, поскольку у некоторых видов в пределах ядра обнаружены медиаторы, отсутствующие у других, что, вероятно, свидетельствует о существовании специфических путей и характеристик системы.
В общем, интерпедункулярное ядро у всех позвоночных характеризуется целым рядом сходных черт: связями с ядрами поводков, гипоталамусом, дорсальным тегментальным ядром, сильной васкуляризацией; наличием нейросекреторных элементов, граничащих с цереброспинальной жидкостью, которые свидетельствуют о его принадлежности к системам мозга, ответственным за организацию целостных поведенческих актов (у высших - к лимбической системе).
Отметим, что упомянутое выше дорсальное тегментальное ядро, nucl. tegmenti dorsalis, или ядро Гуддена, у всех наземных позвоночных характеризуется наличием афферентов от интерпедункулярного
176
ядра и в свою очередь проецируется на моторные ядра краниальных нервов, что позволяет рассматривать его как одно из "выходных" звеньев лимбической системы, обеспечивающих доступ к исполнительным отделам мозга.*
Другой структурой, свойственной мозгу большинства позвоночных, за исключением акул** и круглоротых, является ядро перешейка, nucl. isthmi. Наиболее характерная особенность этой структуры - двусторонние связи с тектумом, которые имеют топологически организованные входы и определенные закономерности становления в онтогенезе. У амфибий, некоторых рептилий и млекопитающих наблюдается сочетанное присутствие в тектуме прямого ипсилатерального входа сетчатки и билатеральных проекций ядра перешейка. В то же время отсутствие ипсилатерального ретинального входа совпадает с отсутствием билатеральных проекций. Показано также, что появление активности нейронов ядра перешейка в ходе эмбрионального развития (у амфибий - в период, предшествующий метаморфозу) приурочено к появлению ипсилатеральных сетчаточных проекций. Данное обстоятельство трудно объяснить однозначно, поскольку волокна упомянутых трактов адресованы разным отделам тектума: ипсилатеральные входы - отделам, находящимся в глубине тектума, а вход ядра перешейка - отделам, лежащим на поверхности тектума, над контрлатеральными ретинальными волокнами. Однако если один из этих компонентов отсутствует, то отсутствует и другой. Такая ситуация характерна для большинства птиц и лучеперых рыб, лишенных ипсилатеральных сетчаточных входов и контрлатеральных проекций ядра перешейка на тектум.
У рептилий ядро перешейка дифференцируют на крупно- и мелкоклеточную части. Первая наиболее развита у крокодилов, ящериц, черепах. У змей, лишенных, по мнению большинства авторов, ядра перешейка, в ростральной части покрышки среднего мозга выделяют nucl. tegmenti posterolateralis, которое на основании сходства связей рассматривают как гомолог (аналог?) ядра перешейка (некоторые авторы описывают ядро перешейка в его обычной локализации).
У птиц различают ядро перешейка, nucl. isthmi, и оптическое ядро перешейка, nucl. isthmoopticus. Первое делят на мелко- и крупноклеточные компоненты и полулунное ядро, раrtes parvo- et magnocellularis et nucl. semilunaris. Для всех компонентов этого комплекса показана конвергенция входов зрительной (от тектума) и слуховой (от ядер латеральной петли)
177
систем. Доказано его участие в реакции сужения зрачка на свет (например, в ответ на звук). Оптическое ядро перешейка описано не у всех видов (например, у Ibis оно замещено полулунным ядром, его дорсальным продолжением). Оно располагается в дорсолатеральной части покрышки и, как обнаружено, принимает участие в центрифугальном контроле чувствительности контрлатеральной сетчатки.
У млекопитающих гомологом ядра перешейка считают-nucl. parabigeminalis, его же рассматривают как гомолог мелкоклеточной части ядра перешейка рептилий и полулунного^ ядра птиц. Это ядро представляет собой небольшую группу клеток, занимающую латеральное положение в среднем мозге, вентрально от медиального коленчатого тела и ручек задних холмов. В ряде случаев в его составе выделяют несколько клеточных групп. Общей чертой всех исследованных в этом отношении видов являются двусторонние связи этого ядра с той частью передних холмов, которая получает ретинальные входы. При этом, если тектальные афференты преимущественно ипсилатеральны, то для обратных связей характерна билатеральность. Учитывая существующую ретинотопию, ряд авторов рассматривает ядро parabigeminalis как сателлитное для передних холмов. Таким образом, nucl. parabigeminalis, подобно ядру перешейка всех позвоночных, оказывается тесно связанным с мезенцефалическим зрительным центром (тектум или передние холмы) и обеспечивает поступление в него информации от контрлатеральной одноименной структуры. Однако, если у большинства позвоночных таким способом, вероятно, обеспечиваются бинокулярные взаимодействия, то у млекопитающих, которые имеют ипсилатеральные ретинальные проекции* возможны и другие функции ядра. Отметим, что у одного вида галаго обнаружены проекции nucl. parabigeminalis в латеральное коленчатое тело, что может быть свойственно и другим животным. Еще одна возможная роль этого ядра - усиление разрешающей способности зрительной системы через активацию истмо-ретинальных проекций.*
Другой структурой, описанной в среднем мозге позвоночных (за исключением круглоротых), является интерколлику-лярное ядро, nucl. intercollicularis, лежащее в центральной области тегментума. Оно получает спинальные входы и проецируется на таламус. У рептилий оно часто рассматривается как место конвергенции акустических и соматических входов. Иногда его включают в состав торуса в качестве перитораль-ной области, regio peritoralis. Вместе с тем, у некоторых животных ядро с таким названием имеет совершенно другие связи.
178
Так, у птиц одноименная структура рассматривается как часть тектума, и ее связи в основном адресованы нижележащим структурам. У акул и скатов интерколликулярное ядро располагается в каудальной части покрышки среднего мозга; оно получает билатеральные входы от тектума и проецируется на спинной мозг, являясь, таким образом, релейной структурой для текто-спинальных связей. Наконец, отметим, что у рептилий название "интерколликулярное ядро" дано двум разным структурам, одна из них, вероятно, опосредует связи стриатума с тектумом.
У млекопитающих выделяют как самостоятельное образовавание центральное серое вещество (ЦСВ) среднего мозга, substantia grisea centralis mesencephali, которое, как полагают, гомологично перивентрикулярной области тектума других позвоночных. Однако связи этих образований значительно различаются. ЦСВ млекопитающих чрезвычайно гетерогенно, содержит несколько типов нейронов, характеризуется разнообразием медиаторов. В его составе выделяют ряд отделов, отличающихся в том числе и по связям. Оно получает многочисленные входы, источники которых локализованы во всех отделах ЦИС. Значительная часть этих структур относится к лимбической системе (в том числе медиальная префронтальная кора, амигдала, гипоталамус, латеральное хабенулярное ядро), а также к звеньям моторных систем (моторная область коры, мозжечок, черная субстанция). Наконец, на уровень ЦСВ проецируются сенсорные структуры: ядро солитарного тракта, главное ядро тройничного нерва, зрительные и слуховые центры среднего мозга, нейроны пластин IV и V спинного мозга, Эфференты адресованы многочисленным структурам, в числе которых лимбические отделы (латеральный гипоталамус, амигдала, фронтальная область коры), парафасцикулярный комплекс таламуса, ядра шва, парабрахиальная и дорсолатеральная области покрышки моста. Многие из этих связей приурочены к определенным областям ЦСВ, однако высокая степень коллатерализации его эфферентных нейронов, а также отсутствие детальных исследований на разных систематических группах не позволяют представить детальную картину организации связей ЦСВ.
Тем не менее, имеющиеся данные позволяют отнести этот отдел мозга млекопитающих к структурам лимбической системы. Так, показано, что его тесные связи с гипоталамусом обеспечивают протекание некоторых фаз репродуктивного поведения. Существует мнение, что ЦСВ является структурой такого уровня, где происходит конвергенция лимбической и моторной систем мозга. Значительное число фактов свидетельствует о причастности ЦСВ к системе эндогенной анальгезии. Это же подтверждают характер поступающей информации от ноцицептивных элементов спинного мозга, наличие опиоидных рецепторов,
179
а также результаты экспериментального изучения влияния ЦСВ на восприятие боли. Однако значительная медиаторная гетерогенность не позволяет пока создать детальную схему взаимодействий этого отдела с другими структурами ноцицептивной системы. Наконец, к настоящему времени накопилось множество данных о его участии в организации видоспецифических вокализаций, которые, как полагают, являются акустическим выражением эмоционального состояния. Это :может вполне согласовываться со сведениями о наличии ноцицептивных входов и афферентов от вышележащих лимбических образований. Таким образом, возможно, что ЦСВ является одним из "выходных" образований лимбической системы, которое преимущественно активирует голосовой аппарат животных и регулирует поступление болевой информации в мозг. Такое сочетание данных представляет интерес и в качестве доказательства справедливости представления о том, что вокализация в процессе эволюции возникает как средство выражения внутреннего состояния (возможно, по принципу смещенной активности) и лишь позднее приобретает коммуникационные функции.
Сложный характер связей и неполнота представлений о роли данного отдела затрудняют проведение сравнительно-анатомического или функционального исследования, поскольку речь идет о структуре, включаемой в состав многокомпонентной лимбической системы млекопитающих, сведения о наличии :и уровне развития которой в мозге других позвоночных отсутствуют.
Таким образом, на уровне среднего мозга у всех позвоночных представлены сенсорные образования зрительной и октаво-латеральной систем. Число моторных образований невелико, и они контролируют сокращение экстраокулярных мышц. Многочисленные связи с исполнительными отделами обеспечиваются на этом уровне надсегментарными (премоторными) образованиями: глубокими слоями тектума, ядром МПП, дорсальным сегментальным ядром, красным ядром, на которых происходит конвергенция входов от сенсорной и лимбической систем, определяя тем самым важную, а в ряде случае решающую роль этого отдела в организации системных поведенческих реакций.
180
*
Отметим, что в составе пучка проходят и восходящие проекции, в; частности соединяющие вестибулярный и глазодвигательный комплексы, а также связи между ядрами III, IV и VI черепномозговых нервов.
*
Латеральное ядро покрышки у
Hydrolagus ранее описано как nucll dorsalis thalami или nucl. rotundus mesencephali.
*
У млекопитающих усложнение лимбической системы приводит и к усложнению связей ядра, но их характер остается прежним: афференты - от лимбических структур, эфференты - к моторным и вегетативным ядрам.
**
В мозге акул ядро с таким названием описано, но кроме сходства в местоположении других данных, свидетельствующих о гомологичности структуры, не имеется.
*
На уровне среднего мозга располагаются и другие структуры, связанные с обработкой зрительной информации. Они входят в состав добавочной зрительной системы и будут, описаны в главе 7, поскольку включают и диенцефальные образования.
