Дорсальную часть продолговатого мозга подразделяют на латерально расположенную соматосенсорную и лежащую медиально висцеросенсорную зоны (рис. 28).
В соматосенсорной зоне различают основное и нисходящее ядра тройничного нерва, а также ядра дорсальных канатиков; эти структуры являются областью краниального представительства соматосенсорной системы.*
Система проекций V черепномозгового нерва имеется уже в мозге круглоротых, где его представительство гомологизируют с нисходящим (спинальным) ядром, nucl. descendens n. trigemini. С развитием системы кожных рецепторов происходит дальнейшее увеличение не только размеров данного ядра, но и области мозга, получающей проекции V нерва. Так, у бесхвостых амфибий значительная часть нисходящего ядра располагается в пределах спинного мозга, фактически замещая желатинозную субстанцию. У челюстных выделяется среднемозговое ядро тройничного нерва, nucl. mesencephalicus n. trigemini (см. гл. 5). У рептилий от спинального отделяется основное (главное) ядро тройничного нерва, nucl. principalis n. trigemini,-составляющее вместе со спинальным единую колонну клеток, которая тянется от уровня вхождения корешка V нерва до ростральной части спинного мозга (рис. 31).
71
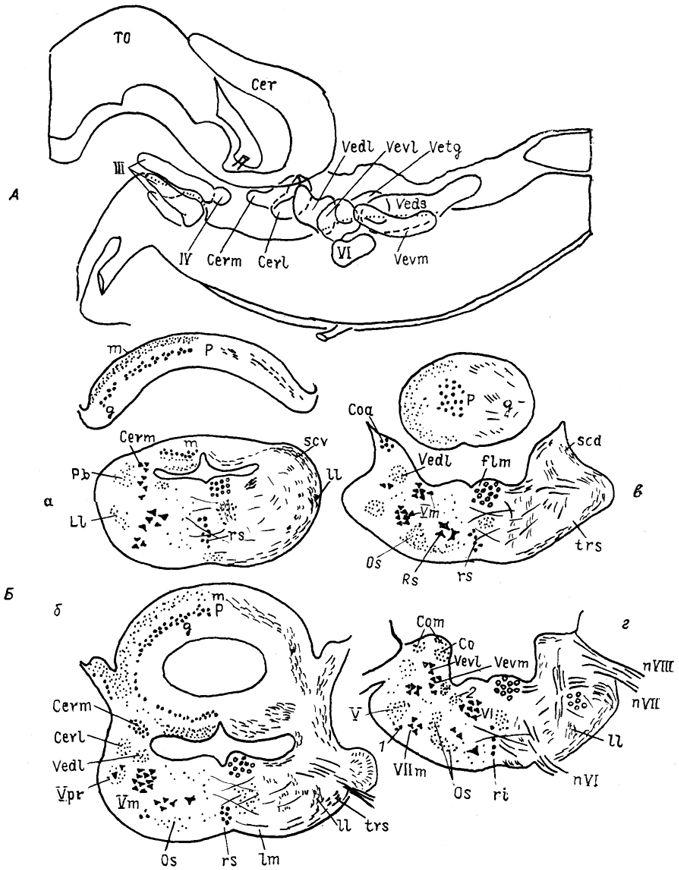
Рис. 31. Строение ромбовидного мозга варана
Varanus exanthematicus на сагиттальном
(А) и фронтальных
(Б) сечениях (Bangma, Donkelaar, 1984).
а-г - срезы в рострокаудальном направлении; 1 - верхнее слюноотделительное ядро, 2 - предлежащее ядро XII нерва.
72
Основное ядро V нерва значительно увеличивается в эволюции, особенно в связи с дифференцировкой сенсорных образований головы. У птиц система тройничного нерва имеет некоторые особенности (рис. 32). Глазничная и максиллярная ветви V нерва приносят информацию от телец Грандри и других тангорецепторов, расположенных в области головы и клюва. Размеры и форма основного ядра значительно варьируют в зависимости от особенностей строения головы. Отсутствие же язычной ветви V нерва, возможно, объясняет наличие входов от других ганглиев: от каменистого (система IX Нерва) к спинальному и главному ядрам, яремного (система X нерва) - к каудальным отделам спинального ядра. Среди млекопитающих особое развитие основного ядра тройничного нерва обнаруживается у однопроходных, грызунов, свиней, что, по-видимому, связано с высокой чувствительностью рострума.
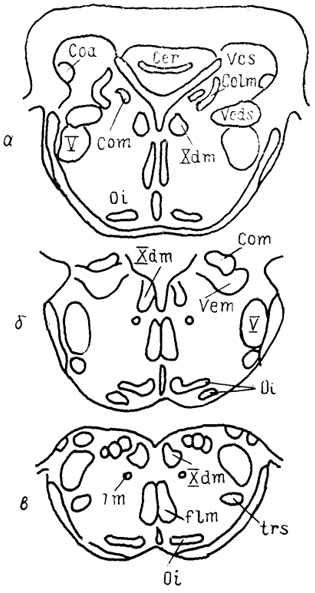
Рис. 32. Схема строения ромбовидного мозга утки
Anas platyrhyncos (Arends e. a., 1984).
а-в - срезы в рострокаудальном направлении.
К числу специализированных черт, возникающих в процессе адаптивной" эволюции, относится иннервация системой тройничного нерва области Якобсоновой ямки - терморецептивного органа, воспринимающего инфракрасное излучение. Он имеется у некоторых змей. В этом случае центральные проекции V нерва состоят из двух компонентов, один из которых подобен имеющимся у всех позвоночных, а другой представлен крупным нисходящим трактом, который заканчивается в отдельном ядре. Эти структуры расположены латерально от "обычного" нисходящего тракта и называются латеральным нисходящим трактом и латеральным нисходящим ядром тройнично-го нерва (рис. 33).
Эфференты ядер тройничного нерва адресованы большому числу образований и в первую очередь моторным ядрам ствола
73
и ретикулярной формации (у акул - спинному мозгу). Часть входов направляется в мозжечок.
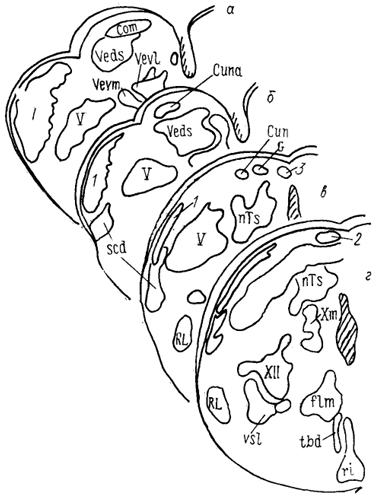
Рис. 33. Ядра каудальной части ромбовидного мозга питона
Python reticulatus (Molenaar, 1977).
а - г - срезы в рострокаудальном направлении. 1 - нисходящее латеральное ядро тройничного нерва, 2 - ядро дорсального канатика, 3 - ядро Бишоффа.
Восходящие проекции с сохранением соматотопии направляются к среднему, промежуточному и конечному мозгу. У всех позвоночных в среднем мозге структурой, получающей тригеминальные входы, является тектум, а у большинства - и тегментум. У млекопитающих наибольшее число входов адресовано вентро-базальному комплексу таламуса, куда они проецируются в виде тройничной петли, lemniscus trigeminus, в составе медиального лемниска, lemniscus medialis, - крупного пучка волокон, связывающих нижележащие отделы с дорсальным таламусом, который объединяет эфференты большого числа соматосенсорных структур. Таламические входы в виде тройничной петли описаны и у рептилий. Вместе с тем у рептилий и птиц обнаруживаются прямые проекции в конечный мозг: эфференты главного ядра восходят в составе квинтофронтального тракта, tr. quintofrontalis, и распределяются в ядрах теленцефалона.
Полагают, что в процессе эволюции позвоночных происходит перераспределение эфферентов тройничного комплекса: у низших их основным источником служит спинальное ядро, а связи приурочены преимущественно к моторным структурам, у высших же возрастает объем проекций к вышележащим отделам, причем основной их источник - главное ядро тройничного нерва. Отметим, однако, что небольшое число билатеральных входов в таламус от нисходящего ядра показано у акул.
Соматосенсорная информация на уровне медуллярного отдела проводится и через ядра дорсальных канатиков, nucll.
74
funiculorum dorsales. Они служат местом окончания путей спинного мозга, восходящих в задних канатиках и в основном не переключающихся на ядрах спинного мозга. У низших позвоночных эти отделы изучены слабо, тем не менее, рядом исследователей обнаружены у актиноптеригий длинные восходящие проекции спинального уровня к каудальной части продолговатого мозга. У многих видов бесхвостых амфибий описаны медиальное и латеральное ядра дорсальных канатиков, nucll. funiculi dorsale medialis et lateralis, первое получает входы от волокон поясничных и грудных сегментов, второе - от цервикального отдела. На этом основании медиальное ядро рассматривается как гомолог нежного ядра, nucl. gracilis, а латеральное - как гомолог клиновидного ядра, nucl. cuneatus, высших позвоночных. Для одного из исследованных видов показаны эфференты этих ядер, формирующие вход в контрлатеральный дорсальный таламус.
Комплекс ядер дорсальных канатиков описан у представителей разных групп рептилий. Так, у черепах он представляет собой группы клеток, рассеянных в дорсальных отделах каудальной части ромбовидного мозга. У питона и других безногих рептилий имеется единая клеточная группа с медиальным и латеральным отделами, которые, возможно, гомологичны нежному и клиновидному ядрам. В этом комплексе обнаруживается соматотопическая организация, подобная таковой у млекопитающих. Срединная группа клеток данного комплекса является местом окончания афферентов области хвоста и гомологизируется с ядром Бишоффа. Наличие добавочного клиновидного ядра, у рептилий отрицается многими авторами, однако, оно описано у питонов как небольшая клеточная группа, состоящая из крупных мультипсляров и занимающая ростральное положение относительно основных ядер комплекса (рис. 33). Эфференты этого ядра адресованы мозжечку. Проекции остальных ядер формируют внутренние дугообразные (аркуатные) волокна и затем продолжаются в виде медиальной петли, восходящей на уровень промежуточного мозга (рис. 31). Точное их окончание в мозге большинства рептилий не выяснено. Для некоторых ящериц доказаны проекции в таламус.
В мозге птиц не всегда возможно разделение ядер задних канатиков на нежное, клиновидное и добавочное клиновидное ядра. Тем не менее этот комплекс формирует бульбо-таламический тракт - гомолог медиальной петли рептилий и части одноименного образования млекопитающих - и путь, адресованный гипоталамусу. Другие эфференты направляются в красное ядро, ретикулярную формацию и небольшая часть - в мозжечок.
Система ядер дорсальных канатиков у всех млекопитающих включает нежное, клиновидное и добавочное клиновидное ядра. Входы к ним сформированы аксонами нейронов спинномозговых
75
ганглиев (и в небольшом количестве - аксонами нейронов задних рогов спинного мозга), проводящими информацию от области задних и передних конечностей (в нежное и клиновидные ядра соответственно). Все ядра дорсальных канатиков взаимосвязаны и получают коллатерали волокон пирамидного тракта, которые терминируют на соме нейронов, в отличие от сенсорных афферентов, оканчивающихся на дендритах.
Состав клиновидного ядра наиболее сложен, оно подразделяется на ростральную и каудальную части. Нейроны каудальной части сгруппированы в вертикально ориентированные колонки, каждая из которых получает афференты определенного дорсального корешка от небольшой по площади рецептивной зоны. При этом обнаружено, что рецепторы разных групп, лежащие в пределах одной рецептивной зоны, проецируются к разным нейронным объединениям. Ростральная часть получает проекции от больших по площади рецептивных поверхностей и здесь обнаруживается перекрытие разных рецептивных зон. Помимо представительства передних конечностей, на уровне клиновидного ядра описаны входы от других сенсорных структур: нисходящего ядра V нерва, вестибулярных и улитковых ядер. Таким образом, здесь имеет место конвергенция разно-модальных входов. Эфференты клиновидного ядра направляются в таламус, причем основным источником этих проекций является каудальная часть ядра, ростральная же формирует проекции и к другим структурам - к тектальной и претектальной областям, к ядрам моста и небольшое число эфферентов - к нижней оливе.
Добавочное (наружное, или латеральное) клиновидное ядро, nucl. cuneatus accessorius, получает входы от сенсорных нейронов, иннервирующих верхнюю часть тела и верхние конечности. Основная масса эфферентов в составе бульбо-мозжечкового тракта направляется к коре мозжечка, небольшая часть - к дорсальному таламусу.
Строение нежного ядра отличается отсутствием выраженных нейронных объединений. Основные входы организованы нейронами, иннервирующими нижние конечности, а эфференты в составе медиальной петли адресуются таламусу.
У млекопитающих часть соматической информации переключается на небольших группах клеток, лежащих вблизи ядер дорсальных канатиков и составляющих ядра X, Y, Z. Сюда, как правило, подходят аксоны спинальных нейронов, принимающих информацию от мышечных рецепторов задних конечностей. Эфференты направляются в таламус и мозжечок.
Таким образом, система тройничного нерва и комплекс ядер дорсальных канатиков представляют собой медуллярный отдел соматосенсорной системы, который обеспечивает проведение информации в вышележащие отделы мозга и характеризуется
76
высоким уровнем соматотопической организации.
Висцеросенсорная зона продолговатого (мозга содержит область проекций VII, IX и X нервов. Активирующие данную область воздействия воспринимаются химическими рецепторами, сосредоточенными у водных позвоночных на поверхности головы, жабрах, плавниках, в глотке, а у наземных - в ротовой полости. Система специализированной хеморецепции обеспечивает кодирование химических стимулов и таким образом обусловливает восприятие качества пищевых веществ (наземные позвоночные) и химических компонентов окружающей среды (водные позвоночные).* Данная система (применительно к водным ее часто называют химической необонятельной, а к наземным- вкусовой) у всех позвоночных построена сходным образом: тела сенсорных нейронов располагаются в ганглиях соответствующих нервов, а их аксоны, формируя одиночный пучок, направляются к сенсорным ядрам продолговатого мозга. У низших позвоночных преимущественно развиты ядра VII и X нервов, nucll. n. facialis et n. vagi, соответственно, а у высших - IX нерва, nucl. n. glossopharingei. При этом у наземных сенсорные части всех трех нервов, сливаясь, образуют единый комплекс - ядро одиночного пучка, или ядро солитарного тракта, nucl. tr. solitarii. В эту систему включается и главное ядро тройничного нерва, куда проецируется импульсация, возникающая при раздражении соматических рецепторов ротовой полости.
Вариации, которые обнаружены в данной системе, обусловлены разного рода адаптациями и особенно значительны на рецепторам уровне. Наиболее многочисленны подобные хеморецептуры у рыб, для которых данный вид чувствительности связан с определением не только наличия съедобных веществ в воде, но и качества воды, пригодности ее для обитания, а также с выбором направления передвижения. У многих амфибий и рептилий количество вкусовых рецепторов невелико, некоторые птицы и змеи их практически лишены. У млекопитающих число вкусовых рецепторов значительно варьирует, однако корреляции с видом пищи и типом питания обнаружить не удастся.
В зависимости от числа и концентрации хеморецепторов разной оказывается и степень развития центральных проекций данной системы.
77
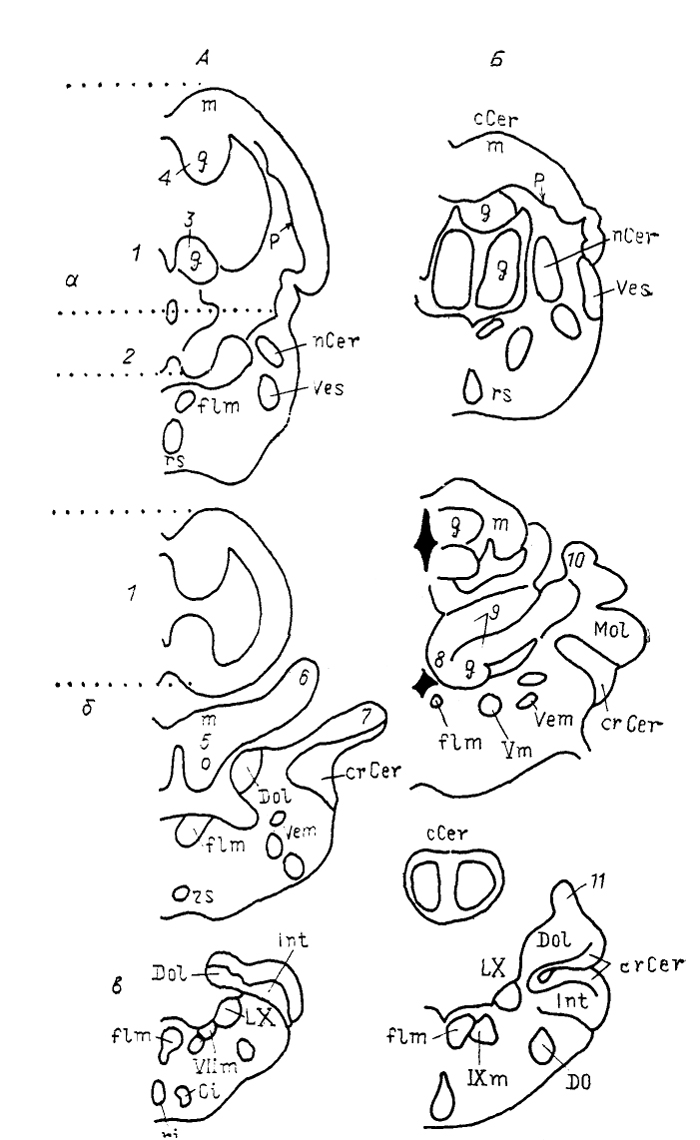
Рис. 34. Ядра ромбовидного мозга акулы
Scyliorhinus canicula (А) и ската
Raja clavata (Б) (Smeets, 1981).
а-в - срезы в рострокаудальном направлении. 1, 2 - дорсальная (1) и вентральная (2) части тела мозжечка; 3, 4 - вентральное (3) и дорсальное (4) гранулярные возвышения; 5-7 - медиальная часть, или нижняя губа (5), верхний (6) и нижний (7) листки латеральной части аурикулы мозжечка; 8 - медиальная гранулярная область; 9-11 - медиальная (9) и латеральная (10) части, дорсальный гранулярный гребень. (11) вестибуло-латеральной доли.
78
В мозге хрящевых рыб висцеросенсорная зона представлена единой клеточной массой, формирующей продольный вырост на латеральной стенке IV желудочка, часто обозначаемой как вагусная доля, lobus vagi (рис. 34, в). Она состоит из мелких клеток и не разделяется на подъядра. Ее эфференты изучены недостаточно полно. Известно, что часть волокон направляется в область перешейка. У одного из видов акул Scyliorhinus здесь описано ядро G. Оно получает входы от вагусной доли и в свою очередь проецируется на спинной мозг. По характеру связей его рассматривают как гомолог вторичного вкусового ядра костных рыб.
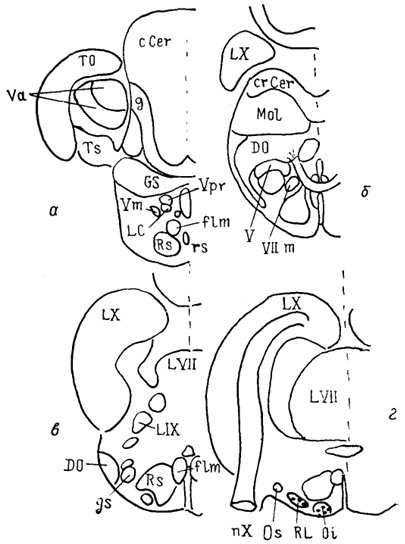
Рис. 35. Ядра ромбовидного мозга представителя костистых рыб
Carassius auratus (Wullimann, Northcutt, 1988).
а-г - срезы в рострокаудальном направлении.
В проекции химической необонятельной системы актиноптеригий прослеживается четкий топологический принцип организации входов: хеморецепторы передней части области рта, поверхности губ иннервируются волокнами VII нерва, задней части рта и жаберных дуг - волокнами IX и X нервов. В зависимости от зоны максимальной концентрации рецепторов изменяется и степень развития центральных отделов. Так, преимущественная локализация рецепторов в полости рта приводит к разрастанию области представительства IX нерва (что имеет место у наземных позвоночных), расположение рецепторов на поверхности соответствует увеличению ядра VII нерва.
Особенно значительны вариации первого центрального отдела у рыб, так как у водных позвоночных функции системы химической необонятельной чувствительности оказываются более широкими, чем у вкусовой системы наземных позвоночных. У рыб, обитающих в мутной воде, или придонных рыб система
79
хоморецепторов имеет не только огромное по площади, но и раздельное представительство в мозге. Так, у p. Carassius вкусовые луковицы, расположенные на поверхности тела, иннервируются системой VII нерва и проецируются в крупное образование, расположенное по средней линии ромбовидного мозга, - лицевую (непарную) долю, lobus facialis (impar) (рис. 35). Информация, воспринимаемая рецепторами, которые сосредоточены в ротовой полости, проецируется в крупные парные вагусные доли, lobi vagi, - центральное представительство X нерва. Аналогичное раздельное представительство внешнИХ и внутренних вкусовых рецепторов обнаруживается и у других рыб, обитающих в сходных условиях (например, у представителей семейства африканских сомов) (рис. 27, 4; 28, в).
Лицевая доля - крупное внутрижелудочковое образование, состоящее из многих типов нейронов. Ее строению свойственны ламинарная организация и топологический характер связей (например, у карповых). У рыб с менее развитыми вкусовыми центрами закономерный характер расположения клеток может отсутствовать.
Вагусные доли у придонных рыб имеют большие размеры и являются самостоятельным отделом, представляющим собой совокупность элементов висцеросенсорной и висцеромоторной зон. Уникальные черты их организации в мозге карповых объясняются, по-видимому, наличием особого небного органа. Вагусные доли у этих животных занимают весь поперечник нижней стенки продолговатого мозга и дифференцированы не менее, чем на 15 слоев. Проекции организованы строго топологически. Помимо сенсорных элементов данный отдел содержит и моторные клетки, иннервирующие мышечные элементы небного органа.
Проекции первичных висцеросенсорных отделов мозга рыб (кроме общих с другими позвоночными) имеют и свои особенности. Так, большой объем волокон от вагусной и лицевой долей направляется в виде крупного пучка - вторичного вкусового тракта, tr. gustatorius secundarius, к особому образованию, также расположенному в медуллярном отделе мозга - :вторичному вкусовому, или висцеральному, ядру, nucl. gustatorius secundarius (рис. 35, а). Оно представлено более или менее выраженной группой клеток, лежащей в области перешейха. Помимо ипсилатеральных входов сюда поступают и контрлатеральные проекции, после перекреста в комиссуре ядра, а также некоторое количество первичных входов. У видов, для которых данная сенсорная система является ведущей, вторичный вкусовой тракт представляет собой один из крупнейших путей продолговатого мозга, обеспечивающих строго топологически организованное проведение сенсорной информации. У многих актиноптеригий данное ядро имеет сложную структурную организацию и состоит из многочисленных нейронных кластеров.
Эфференты вторичного вкусового ядра изучены недостаточно: имеются сведения о его значительных комиссуральных связях, а также о существовании эфферентов к ретикулярной формации
80
и мозжечку. Однако наиболее выраженными оказываются связи этого ядра с каудалъной областью гипоталамуса. Данное ядро формирует и нисходящие связи с первичными вкусовыми центрами.
У амфибий, подобно другим наземным позвоночным, обнаруживается единое ядро солитарного тракта. Его эфференты адресованы вторичному вкусовому ядру, лежащему в области перешейка.
У высших позвоночных в составе ядра солитарного тракта выделяют в ростро-каудальном направлении промежуточное ядро, nucl. interstitialis, нисходящее, nucl. descendens, и комиссуральное, nucl. commissuralis n. vagi, ядра, соответствующие распределению подходящих афферентов VII, IX и X нервов соответственно. Другие афференты происходят из латерального гипоталамуса, голубого пятна, теленцефалона (у млекопитающих - новой коры). Эфференты адресованы моторным ядрам одноименных черепномозговых нервов и ядру подъязычного нерва. Часть проекций направлена к вышележащим центрам данной сенсорной системы: таламическим ядрам, гипоталамусу, центральному ядру миндалины (у млекопитающих), а также ядрам мостового уровня (у рептилий - к области перешейка). Последние получают основной объем эфферентов ядра солитарного тракта. У птиц и млекопитающих к ним относятся дорсальное и вентральное парабрахиальные ядра, лежащие соответственно дорсолатерально и вентромедиально от верхних ножек мозжечка. У млекопитающих к этому комплексу относится и ядро Колликера - Фузе, расположенное вентролатерально от парабрахиалышх ядер. У рептилий это вторичное вкусовое ядро, или парабрахиальная область, как полагают, гомологичная дорсомедиальной части парабрахиального комплексами ядру Колликера-Фузе млекопитающих (рис. 31, Б, а).
Афферентами вторичных вкусовых центров, помимо входов от ядра солитарного тракта, являются проекции от гипоталамуса, миндалины, двойного ядра. Их эфференты адресованы двойному ядру, ретикулярной формации, спинному мозгу. Кроме того, у млекопитающих вентральное парабрахиальное ядро проецируется в таламус, дорсальное - во фронтальную кору, гипоталамус и миндалину. Две последние структуры принимают входы и от ядра Колликера-Фузе.
У разных млекопитающих несколько отличается характер висцеросенсорных центров. Так, если у грызунов входы в ядро солитарного тракта достаточно четко разделены, то у многих приматов обнаруживаются значительные области перекрытий афферентов, относящихся к разным нервам. Кроме того, у приматов основным уровнем, получающим эфференты ядра солитарного тракта, является таламус, а не парабрахиальные ядра. Последние же в меньшей степени формируют входы в таламус
81
и гипоталамус и в основном адресуют проекции в миндалевидный комплекс. Таким образом, у высших млекопитающих обнаруживаются восходящие проекции с меньшим числом переключений.
Многочисленные связи медуллярных сенсорных ядер данной системы с моторными и вегетативными центрами обеспечивают протекание таких приспособительных реакций, как глотание, жевание, слюноотделение, рвотный рефлекс. Восходящие проекции к среднему мозгу и мозжечку (у низших позвоночных), гипоталамусу и таламусу (у высших) обусловливают сложные формы пищевого и питьевого поведения.
Область центральных проекций висцеросенсорных нервов является также одним из звеньев висцеральной сенсорной системы, которая опосредует передачу информации из внутренней среды организма от хеморецепторов внутренних органов и участвует в организации регуляторных процессов.
Область представительства интерорецепторов у низших позвоночных не отделена от вкусового представительства (например, вагусная доля рыб), у наземных позвоночных она приурочена к каудальной части ядра солитарного тракта, а ее проекции сосредоточены преимущественно в латеральной части парабрахиального комплекса. Последний проводит информацию к моторным и вегетативным ядрам черепномозговых нервов, а также в гипоталамус и лимбические отделы промежуточного и конечного мозга, обеспечивая протекание интероцептивных рефлекторных реакций и создание разного рода мотивацион-ных состояний.
Акустико-латеральная область составляет значительную часть дорсолатеральных отделов продолговатого мозга. Она рассматривается в качестве деривата соматической сенсорной: колонны на основании как своего положения в мозге, так и происхождения рецепторов данной системы из эктодермальных. производных - плакод. Данная область мозга является зоной-представительства октаво-латеральной системы, объединяющей органы специализированной механорецепции и их центральные проекции. Октаво-латеральная система на периферии включает в себя сообщающиеся с внешней средой органы боковой линии и изолированный от внешней среды отдел-лабиринт. Первые обнаруживаются только у водных позвоночных и обеспечивают поступление информации о перемещениях как самого животного, так и водной среды. У некоторых групп низших позвоночных имеются специализированные органы боковой линии, чувствительные к электрическим воздействиям (миноги, некоторые рыбы, хвостатые амфибии). Электрорецепторы таких органов адаптированы к восприятию градиента электрического поля и таким образом опосредуют реакции животного как на электрические поля других особей, так и на изменения собственного электрического поля. Октавная подсистема получает
82
информацию от механорецепторов, лежащих в лабиринте. Лабиринт содержит два функционально различных отдела - вестибулярный аппарат, который обеспечивает восприятие перемещений тела и является органом гравитации, и внутреннее ухо - звуковоспринимающий отдел (последний как самостоятельный отдел у низших позвоночных отсутствует, а его функции выполняют элементы вестибулярного аппарата).
Октавная подсистема обнаруживается у всех позвоночных, органы боковой линии - у всех анамний в личиночной стадии и у большинства взрослых анамний. Менее широко представлена электрорецептивная часть: она отсутствует у голостей, большинства телеостей и некоторых амфибий.
Сходство в структуре и характере развития рецепторов разных отделов октаво-латеральной системы привело к формированию представления об общности их происхождения. Ранее полагали, что механорецепторы канальных органов боковой линии дают начало электрорецепторам и рецепторам внутреннего уха, причем среди последних раньше возникают вестибулярные рецепторы. Слуховая рецепция появляется лишь у переходных к амфибиям форм с образованием передающих аппаратов среднего уха и рецепторов, способных отвечать на звуки, распространяющиеся по воздуху. Переход от водной к наземной среде обитания у многих амфибий знаменуется утратой органов боковой линии, а проекции новой системы адресуются той же области продолговатого мозга, изменяя ее функции. Однако, современные морфологические и физиологические данные свидетельствуют о самостоятельности и параллелизме в развитии каждого компонента октаво-латеральной системы. Иными словами, каждый из них, появившись рано и одновременно в эволюции позвоночных, является "базовым" отделом продолговатого мозга. Изменения, которые затем происходят в каждой из субсистем, включают адаптивные преобразования и центрального представительства (в том числе редукцию центральных отделов системы органов боковой линии с исчезновением соответствующих рецепторов).
Каждая часть октаво-латеральной системы имеет собственные пути проведения, представленные краниальными нервами, а также специализированную область в пределах акустико-латеральной зоны продолговатого мозга.
У животных, обладающих органами боковой линии, в организации путей проведения обнаруживается ряд общих черт. Так, у круглоротых и рыб информация от рецепторного уровня поступает в мозг по трем парам нервов: дорсальному и вентральному передним нервам боковой линии, nn. lineae lateralis anterior dorsalis et ventralis, и заднему нерву, n. lineae lateralis posterior. Чаще всего их рассматривают как специализированные отделы лицевого (передний нерв) и блуждающего (задний нерв) черепномозговых нервов; однако некоторые авторы отстаивают их самостоятельное происхождение.
Центры представительства органов боковой линии расположены в дорсальной и промежуточной зонах акустико-латеральной области и покрыты снаружи весьма характерным образованием - мозжечковым гребнем, crista cerebelli, который является продолжением молекулярного слоя мозжечка (рис. 34, 35).
83
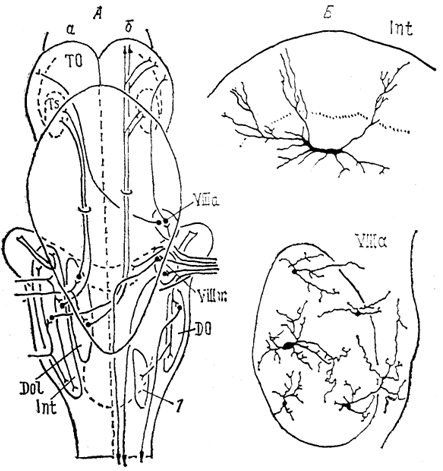
Рис. 36. Схема связей
(А) и нейронный состав
(Б) ядер октаво-латеральной области хрящевых рыб (Smeets e. а., 1983).
а, б - связи ядер органов боковой линии (а), VIII нерва (б). 1 - ядро бокового канатика.
В пределах мозжечкового гребня формируются контакты нейронов центров боковой линии с параллельными волокнами и аксонами звездчатых клеток мозжечка.
Система связей центров боковой линии изучена недостаточно полно. Показано, что крупные клетки, входящие в их состав и получающие первичные афференты, в свою очередь проецируются на элементы ретикулярной формации. Восходящие проекции в средний мозг обнаружены у пластиножаберных и некоторых телеостей (рис. 36, А).
Степень и характер развития области представительства органов боковой линии имеет ряд отличий у позвоночных разных групп. Так, у миног и пластиножаберных, а также у некоторых костных рыб (хондростей, двоякодышащих и многоперовых) в ее пределах выделяют дорсальное и медиальное ядра, nucll. dorsalis et medialis, иначе переднюю и заднюю доли боковой линии, lobi lineae lateralis anterior et posterior (рис. 29, 34). Полагают, что у пластиножаберных имеется вход В торус среднего мозга через примитивную систему латеральной петли. Однако, по мнению других исследователей, эфференты
84
дорсального и медиального ядер направляются контрлатерально в субтектальные области.
У миксин, голостей и телеостей проекции органов боковой линии адресованы медиальному ядру акустико-латеральной зоны. У телеостей оно состоит из дорсальной и медиальной частей. Первая более дифференцирована, дендриты ее нейронов проникают в мозжечковый гребень и получают входы от заднего нерва боковой линии (от рецепторов туловищного отдела). Медиальная часть не имеет четких границ и связана с представительством рецепторов головы, иннервируемых передним нервом боковой линии. Дорсальное ядро у этих животных отсутствует (рис. 35, б).
Возможным объяснением подобных различий в организации первого центрального отдела системы органов боковой линии является специализация ее в связи с электрорецепцией. Действительно, среди водных позвоночных обнаружены виды, чувствительные к электрическим воздействиям и воспринимающие низкочастотные и низкоамплитудные сигналы, которые распространены в водной среде. Электрочувствительными являются миноги, некоторые представители рыб и хвостатых амфибий. Отдельные виды рыб способны не только воспринимать, но и генерировать электрические разряды (так называемые электрогенные рыбы). Подобного рода особенности, а также происхождение элементов электросенсорных систем обусловили различия в характере организации центральных звеньев органов боковой линии.
Полагают, что дорсальное ядро октаво-латеральной системы является структурой, специфически связанной с электрорецепцией. Об этом же свидетельствует его присутствие в мозге электрочувствительных амфибий. Вместе с тем электрочувствительные костистые рыбы (клюворылые, гимнотовидные, сомообразные и араванообразные) лишены дорсального ядра. У них информация от электрорецепторов по системе нервов боковой линии достигает особой, электросенсорной, области, которая занимает латеральную часть акустико-латеральной зоны продолговатого мозга (рис. 28, д, е). В этой области обнаруживается ряд структурных изменений по сравнению с медиальной частью, куда проецируются механорецепторы органов боковой линии.
Так, под глубоким слоем мозжечкового гребня формируется плексиморфный слой, в котором располагаются дендриты клеток, подобных клеткам Пуркинье мозжечка. На них, а также на телах округлых клеток этого слоя заканчиваются аксоны первичных сенсорных нейронов. Крупные клетки являются проекционными, направляющими аксоны к вышележащим отделам, округлые клетки - комиссуральными.
У рыб, способных генерировать электрические сигналы, обнаруживается дальнейшая специализация области центрального представительства органов боковой линии. В медуллярном отделе значительно утолщается молекулярный слой, зона крупных клеток дифференцируется на пять самостоятельных слоев, число клеточных типов возрастает до десяти. При этом разные типы клеток получают информацию от разных видов рецепторов, а эффекторные элементы медуллярного уровня проецируются в разные отделы торуса среднего мозга (рис. 37). В ряде случаев у электрогенных рыб гипертрофированными оказываются система латеральной петли и торус среднего мозга (рис. 38).
Считается, что электрорецепция в эволюции появляется сравнительно рано - у животных, давших начало миногам и хрящевым рыбам. Однако, хотя она сохраняется во многих линиях позвоночных, включая предков наземных, в более молодых группах - у всех ныне живущих голостей и большинства
85
телеостей - она, по-видимому, была утрачена, что сопровождалось исчезновением дорсального ядра октаво-латеральной области. В настоящее время лишь у 1% костистых рыб имеются органы электрорецепции. При этом и
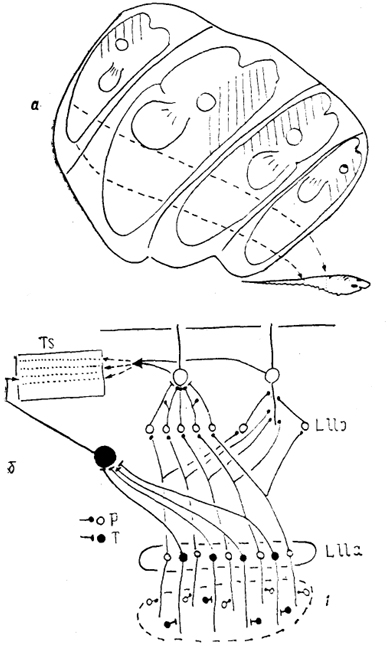
Рис, 37. Схема организации проекций электрорецепторов в мозге
Gymnoti-formis (Maler e. a., 1981; Heiligenberg, Dye, 1982).
а - распределение входов в заднюю долю органов боковой линии от рецепторов разных отделов тела, б - представительство афферентов от двух типов бугорковых рецепторов (Р и Т) в стволе мозга. 1 - рецептивная поверхность.
рецепторная часть и область центральных проекций электросенсорной системы значительно отличаются от таковых у видов "основной" линии, что может свидетельствовать о ее независимом и позднем появлении в эволюции.
Таким образом, медуллярное представительство органов боковой линии у разных групп рыб (рыбы, не обладающие электрорецепцией, костистые рыбы с электросенсорной системой, электросенсорные рыбы, не относящиеся к костистым, и электрогенные
86
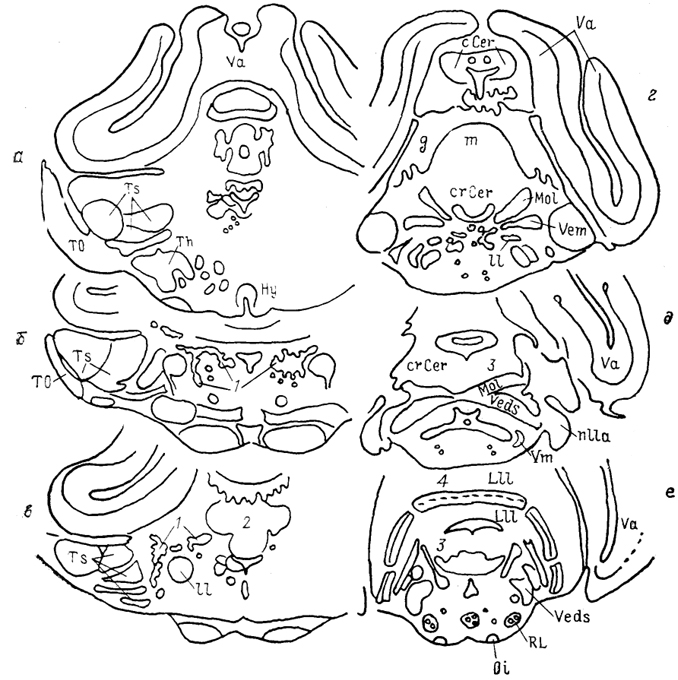
Рис. 38. Ядра ствола мозга представителя мормирид
Gnathonemus petersi (Meek e. а., 1986),
a - e - срезы в рострокаудальном направлении. 1 - латеральное ядро заслонки мозжечка; 2 - гранулярное возвышение, 3, 4 - области проекций механо- (3) и электрорецепторов (4) в зоне представительства органов боковой линии.
рыбы) организовано по-разному, а электрорецепция и связанные с нею мозговые образования костистых рыб не гомологичны таковым у других групп животных.
У всех амфибий система органов боковой линии присутствует, по крайней мере в личиночной стадии. В периферическом отделе она включает механо-, а у некоторых и электрорецепторы. Центральная организация медуллярного представительства подобна той, которой обладают рыбы, не относящиеся к костистым. Ядра октаво-латеральной системы амфибий занимают перивентрикулярное положение.
87
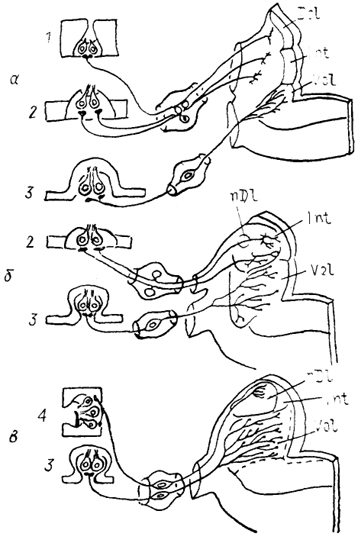
Рис. 39. Октаво-латеральная область продолговатого мозга амфибий (Fritzsch e. а., 1985).
а - электрочувствительные хвостатые амфибии, б - хвостатые и личинки бесхвостых, в - взрослые бесхвостые амфибии. 1-4 - рецепторы октаво-латеральной системы: электрорецепторы (1), механорецепторы органов боковой линии (2), вестибулярные (3), акустические (4).
У личинок всех амфибий и взрослых особей водных хвостатых амфибий передний нерв боковой линии имеет несколько корешков. Дорсальный корешок проводит информацию в область продолговатого мозга, названную дорсальным островком Кингсбери, или дорсальным ядром октаво-латеральной области, nucl. dorsalis areae octavolateralis. На основании томографического сходства и наличия входов от электрорецепторов его гомологизируют с дорсальным ядром примитивных электрорецептивных рыб. В зону, лежащую вентрально от него, входит пучок, проводящий информацию от механорецепторов органов боковой линии. Этот отдел-гомолог медиального ядра октаво-латеральной области рыб - носит название промежуточного ядра, nucl. intermedius areae octavolateralis (изначально описано как каудальное ядро), и у бесхвостых амфибий перед метаморфозом редуцируется до маленькой клеточной группы. По имеющимся данным оно получает небольшое число входов от вестибулорецепторов (рис. 39). У личинок бесхвостых амфибий имеется дорсальный островок Кингсбери, однако у взрослых особей, даже обитающих в воде, его нет, что связывают с отсутствием органов электрорецепции. Органы боковой линии, сохраняющиеся у некоторых бесхвостых амфибий, проецируются в промежуточное (медиальное) ядро. Восходящие связи медуллярного центра направляются в средний мозг. У некоторых безногих амфибий сохраняются и механо- и электросенсорные органы боковой линии, однако их центральные проекции недостаточно изучены.
Таким образом, при специализации органов боковой линии,
88
каждая из рецепторных систем раздельно представлена на медуллярном уровне, причем области представительства топологически четко организованы и часто имеют ламинарный характер строения.
Акустическая зона (октавная - у низших) продолговатого мозга включает в себя два комплекса ядер: ядра, расположенные дорсолатерально и связанные с обработкой информации об акустических сигналах и лежащие вентромедиально вестибулярные ядра. Обе группы составляют область представительства VIII черепномозгового нерва,* n. vestibulocochlearis, и занимают вентральное положение относительно центров боковой линии.
В мозге круглоротых система VIII нерва проецируется в вентральную часть октаво-латеральной области (рис. 29). В ее пределах некоторые авторы выделяют три ядра (дорсальное, медиальное, тангенциальное), другие - два (медиальное и заднее), но большинство склоняется к мысли о гомологичноети всей этой области вентральному ядру октаво-латеральной области рыб. Эфференты этого отдела направляются в спинной мозг и в составе медиального продольного пучка - к глазодвигательному комплексу. Имеются сведения о наличии проекций в торус, ядро задней комиссуры и диенцефальную область. Часть входов адресована ретикулярной формации.
В мозге рыб волокна VIII иерва проецируются в вентральную зону октаво-латеральной области продолговатого мозга. Конкретное описание ее разными авторами проводится по-разному и вопросы гомологизации ядер октавного комплекса далеко не разрешены.
Полагают, что вентральное ядро, получающее в мозге круглоротых входы от вестибулорецепторов и проецирующееся на спинной мозг, сохраняется и в мозге рыб (рис. 29, 35, 36). При этом происходит увеличение количества и отчасти размеров составляющих его элементов. У многих групп рыб (хрящевых, саркоптеригий и актиноптеригий за исключением телеостей) описывают четыре ядра системы VIII нерва - переднее, крупноклеточное,** нисходящее и заднее вестибулярные. У костистых рыб появляются дополнительные ядра этого комплекса: ядро Дейтерса и тангенциальное.
Эфференты октавной системы адресуются многим отделам мозга, в ,их числе вышележащие образования - торус, мозжечок, а также моторные ядра ствола. Связи с моторными ядрами образуют особый пучок, tr. octavomotorius, волокна которого проходят в составе иней- и контрлатерального медиальных продольных пучков и формируют восходящую и нисходящую
89
ветви. Восходящая ветвь направляется к ядрам III и IV нервов, нисходящая - к ядру VI нерва и мотонейронам спинного мозга. Волокна, идущие к спинному мозгу, у костистых рыб могут проходить параллельно медиальному продольному пучку. Среди них выделяют перекрещенный латеральный октаво-спинальный тракт, который, как полагают, берет начало от тангенциального ядра, и прямой медиальный октаво-спинальный тракт, идущий от крупноклеточного вестибулярного ядра.
Значительное число эфферентов после перекреста восходит рострально и направляется в торус среднего мозга. Эта система носит название латеральной петли, lemniscus lateralis, или .латерального продольного пучка, fasc. longitudinalis lateralis, и хорошо выражена у всех лучеперых рыб. Ее волокна, помимо торуса, адресуются латеральному ядру заслонки мозжечка, а у костистых рыб - tectum opticum.
Эфференты октавной части хрящевых рыб изучены недостаточно подробно. Имеются сведения о наличии проекций к спинному мозгу и мозжечку (рис. 36, А). У голостей и костистых рыб показано широкое и топографически организованное представительство разных участков вестибулярных ядер в различных отделах мозжечка.
Комиссуральные связи октавной области составлены 'внутренними дугообразными волокнами, которые проходят в области тегментума вентрально от медиального продольного пучка.
Для всех групп рыб обнаружена эфферентная иннервация октавной части. Источники связей локализованы в пределах ретикулярной формации и составляют "октаво-латеральное эфферентное ядро". У костистых рыб описана также дополнительная группа клеток, расположенная в промежуточном мозге.
В мозге амфибий основные входы волокон VIII нерва адресованы вентральному ядру акустико-латеральной области, а их коллатерали (у хвостатых и личинок бесхвостых) - латеральной части промежуточного ядра.* Кроме того, имеются входы и в латеральную зону ретикулярной формации. В составе вентрального вестибулярного ядра у разных видов описано от двух до пяти отделов (рис. 30). Его эфференты образуют октаво-моторный тракт, а само ядро (или его часть) рассматривается как гомолог ядра Дейтерса. Эфференты вестибулярных ядер амфибий формируют и нисходящие проекции к рецепторам вестибулярного аппарата.
В мозге рептилий вестибулорецепторы проецируются в крупноклеточное ядро, nucl. vestibularis magnocellularis, - возможный гомолог ядра Дейтерса - и лежащее латерально тангенциальное ядро, nucl. tangentialis. У многих рептилий, однако,
90
имеется более сложное развитие комплекса и в его составе выделяют до шести ядер (рис. 31). Крупноклеточное ядро (вентролатеральное вестибулярное по другой классификации) я его основной путь - латеральный вестибуло-спинальный тракт - достигают значительного развития у четвероногих рептилий, у безногих же особенно велико тангенциальное ядро.
Нейроны вестибулярного комплекса рептилий формируют два нисходящих тракта - латеральный и медиальный вестибуло-спинальные, trr. vestibulospinalis lateralis et medialis. Первый берет начало от крупноклеточной части вентролатерального и тангенциального ядер и идет ипсилатерально. Второй начинается от контрлатеральных вентромедиального, нисходящего, вентролатерального и от ипсилатерального дорсолатерального ядер. Восходящие проекции адресованы структурам среднего мозга (контрлатеральным ядрам III и IV нервов) и проходят в составе медиального продольного пучка. В ряде случаев доказано, что как восходящая, так и нисходящая ветви могут образовываться одним и тем же нейроном в результате бифуркации аксона. Такие клетки описаны в медиальном и дорсолатеральном вестибулярных ядрах.
В мозге птиц ядра вестибулярного комплекса сохраняют свойственные им состав и связи. Помимо них выделяют и дополнительные ядра - дорсомедиальное и верхнее вестибулярные (рис. 32). Эфференты вестибулярного комплекса у рептилий и птиц оказываются сходными и отличаются по степени выраженности. Основные проекции адресованы мозжечку, моторным ядрам черепномозговых нервов, ретикулярной формации, глазодвигательному комплексу. Имеются и билатеральные входы к рецепторным отделам. Среди вестибуло-спинальных лроекций выделяют латеральный и медиальный тракты. Первый проходит в латеральных канатиках, а второй - в латеральном отделе вентральных канатиков. Кроме того, по системе дугообразных волокон проходят связи с контрлатеральным вестибулярным комплексом. Помимо входов от нейронов ганглия Скарпа в вестибулярный комплекс рептилий и птиц поступают волокна спинно-вестибулярного тракта.
В вестибулярном комплексе млекопитающих значительно редуцируется тангенциальное ядро, а в составе латерального ядра обнаруживаются как крупные, так и мелкие нейроны. Каудальным продолжением мелкоклеточного отдела ядра Дейтерса является нижнее, или нисходящее, ядро. Медиальное •ядро хорошо развито и его гомологи обнаруживаются в мозге рептилий и птиц. Кроме названных в составе вестибулярного комплекса представлено верхнее, или ядро Бехтерева, а также группы клеток X, Y, Z. Входы от нейронов ганглия Скарпа организованы таким образом, что информация от рецепторов полукружных каналов адресуется верхнему и медиальному ядрам, нижнее и латеральное получают входы от рецепторов саккулуса
91
и утрикулуса. Активация рецепторов саккулуса, кроме того, приводит к изменению активности нейронов группы Y, утрикулуса - медиального вестибулярного ядра. Помимо первичных сенсорных входов ядра комплекса получают проекции от разных отделов ретикулярной формации, мозжечка, спинного мозга. Весьма сложными оказываются связи между ядрами комплекса. Комиссуральные связи опосредуются в основном нейронами группы Y.
Эфференты вестибулярных ядер направляются к моторным структурам, мозжечку, ретикулярной формации в составе медиального продольного пучка, вестибуло-спинального и вестибуло-мозжечкового трактов. Часть волокон адресуется ядрам глазодвигательного комплекса (III, IV и VI черепномозговых нервов). Нейроны латерального вестибулярного ядра осуществляют иннервацию рецепториого отдела.
У млекопитающих система проекций вестибулярных ядер значительно усложняется в связи с развитием тонких зрительно-моторных координации. При этом остается открытым вопрос о наличии и функциональной роли проекций вестибулярной системы, восходящих к таламо-теленцефальному уровню.
В целом у позвоночных обнаруживается удивительное сходство в характере связей вестибулярной системы: многочисленные проекции к исполнительным отделам мозга определяют решающую роль данной сенсорной системы во взаимодействии со зрительной, проприоцептивной и соматической системами и обеспечивают осуществление позных рефлексов и ориентацию в трехмерном пространстве.
Наиболее сложной, по-видимому, является проблема эволюционного становления акустической части октаво-латеральной системы, здесь остаются нерешенными вопросы как возникновения, так и гомологизации слуховых центров. Так, неясно, имеют ли бесчелюстные в дополнение к вестибулярным слуховые рецепторы и соответствующие отделы в продолговатом мозге. Считается, что рыбы уже обладают слуховыми рецепторами внутреннего уха, однако отдельного центрального их представительства не обнаружено. Вместе с тем для одной группы рыб показано, что наряду с четко организованными входами вестибулорецепторов в определенные области октавного комплекса проецируются и рецепторы саккулуса, возможно связанные с восприятием акустических стимулов. Это позволяет предположить, что, по крайней мере, некоторые группы рыб, подобно тетраподам, имеют раздельные акустические и вестибулярные области продолговатого мозга и соответственно ветви VIII нерва.* Однако вряд ли это является правилом для организации
92
слуховой системы всех рыб, поскольку рецепторы внутреннего уха у них оказываются гетеромодальными и могут быть активированы как акустическими, так и гравитационными воздействиями. Более того, у разных рыб функцию восприятия акустических раздражителей выполняют разные периферические отделы октавной системы.
Особенно сложен вопрос об организации слуховых центров и их происхождении у амфибий, поскольку именно на этом уровне появляются четка дифференцированные мозговые структуры данной системы. Согласно хорошо известной гипотезе О. Ларселла у амфибий с исчезновением механорецептивных органов боковой линии промежуточное (медиальное) ядро становится центральным представительством слуховой, системы, преобразуясь в кохлеарное, или улитковое, ядро. Однако многочисленные исследования последних лет убедительно опровергли это мнение. Так, у бесхвостых амфибий акустические входы проецируются как в промежуточное, так и в дорсолатеральное ядро октаво-латеральной области продолговатого мозга (рис. 39). Последнее появляется непосредственно перед метаморфозом и становится основным медуллярным отделом слуховой системы. На месте промежуточного ядра у бесхвостых амфибий остается небольшая группа клеток, которая получает коллатерали слухового нерва. У хвостатых амфибий дорсолатеральное ядро отсутствует и акустический вход адресуется промежуточному ядру. Вместе с тем существует множество вариаций этой схемы. Так, у некоторых хвостатых амфибий, развивающихся без метаморфоза, промежуточное ядро не обнаружено. У тех безногих амфибий, у которых доминирует акустический вход, специфические слуховые образования отсутствуют вовсе. Все это позволило ряду авторов высказать предположение о многообразии путей становления слуховой системы в группе амфибий и, таким образом, допустить возможность параллельного развития слуховых образований в разных группах земноводных.
Представляется неясным происхождение акустического центра бесхвостых амфибий - дорсолатерального ядра (оно ранее описано под разными названиями, наиболее известное из которых "дорсальное крупноклеточное ядро". Гипотеза о происхождении его из медиального (механорецептивного в системе органов боковой линии) ядра рыб опровергается, по-видимому, наличием и промежуточного, и дорсолатерального ядер одновременно у тех бесхвостых амфибий, у которых во взрослом состоянии сохраняются органы боковой линии.
Другая линия доказательств независимости возникновения акустического ядра от редукции органов боковой линии проводится исходя из различий в характере эфферентных проекций этих систем. Проекции медуллярных центров адресованы разным отделам торуса, прямые связи слухового ядра с тектумом отсутствуют, наконец, в пределах продолговатого мозга бесхвостых амфибий формируется новое скопление клеток, получающее входы только от слухового ядра,-верхняя олива, oliva superior (рис. 30, 5). Не получает подтверждений и представление о развитии акустического центра из дорсального ядра октаво-латеральной области. Действительно, потеря электрорецептивных
93
органов и появление дорсолатерального ядра, занимающего положение, идентичное положению дорсального (электросенсорного) ядра, совпадают во времени. Исходя из этого было высказано мнение о трансформации данной структуры. Однако изучение генеза нейронов слухового ядра опровергает эти предположения: его источники, по-видимому, локализуются и в вентральной, и в дорсальной частях акустико-латеральной области. Последняя представлена пролиферативной зоной, которая после утраты дорсального ядра дает начало клеткам акустического ядра. Вентральная же, вероятно, гомологична области, занимаемой в мозге рыб вентральным ядром. Об этом свидетельствует сходный характер связей.
Эфференты дорсолатерального ядра направляются к контрлатеральному одноименному образованию, в торус среднего мозга и билатерально в верхнюю оливу, которая располагается, в пределах медуллярной ретикулярной формации и в оформленном виде появляется у личинок бесхвостых амфибий перед, метаморфозом. Слуховое ядро составляет основной объем ее афферентов. Эфференты верхней оливы, а также некоторое число волокон слухового ядра составляют латеральный бульбо-тектальный тракт, или латеральную петлю, и адресуются торусу. Рассеянные среди волокон клеточные тела рассматриваются как предшественники ядер латеральной петли высших, nucll. lemnisci lateralis.
В мозге разных рептилий акустический комплекс обнаруживает значительное число вариаций строения. Так, у черепах, многих ящериц, амфисбен в составе этой области выделяют два ядра, нейроны которых получают входы слуховой ветви-VIII нерва. Ядра имеют разные названия, чаще всего их обозначают как переднее и заднее кохлеарные (улитковые), nucll. cochlearis anterior et posterior. У змей описана лишь одна клеточная группа. Значительно сложнее устроен кохлеарный комплекс у некоторых ящериц (варан, хамелеон, игуана, агама), и особенно у крокодилов. У последних состав и характер связей сходны с имеющимися у птиц. У названных рептилий-в пределах переднего гипертрофированного ядра выделяют ростральное, угловое и дорсальное крупноклеточное кохлеарные ядра, nucl. cochlearis rostralis, angularis et dorsalis magnocellularis соответственно (рис. 31). Полагают, что дорсальное крупноклеточное ядро рептилий - гомолог акустического - ядра амфибий и вентрального кохлеарного ядра млекопитающих, угловое же, не имеющее предшественников в мозге бесхвостых амфибий, превращается у млекопитающих в дорсальное кохлеарное ядро. Эфференты дорсального крупноклеточного ядра направляются к ламинарному ядру, которое рассматривается как гомолог медиальной верхней оливы и, возможно, ядер трапециевидного тела. Восходящие проекции всех слуховых
94
ядер направляются к верхней оливе и в слуховые центры среднего мозга.*
В мозге птиц слуховые центры медуллярного отдела занимают большой объем и имеют сложное строение (рис. 32). Ядра кохлеарного комплекса состоят из дорсального и вентрального ядер. Они рассматриваются как гомологи соответственно углового и крупноклеточного ядер рептилий. От вентрального ядра проекции адресуются ламинарному ядру, nucl. cochlearis laminaris, которое на основании сходства связей считают гомологичным медиальлой верхней оливе млекопитающих-Нейроны углового и ламинарного ядер дают начало крупному пучку волокон (возможно, трапециевидному телу), направленному контрлатерально и передающему информацию верхней оливе и ядрам латеральной петли противоположной части мозга. Часть волокон восходит к среднему мозгу. Наконец, некоторая часть проекций кохлеарных ядер птиц адресована ретикулярной формации и моторным ядрам черепномозговых нервов.
Описание комплекса ядер латеральной петли у птиц достаточно противоречиво. У видов с развитой слуховой системой большинство авторов выделяет в его составе верхнюю оливу, вентральную часть ядер латеральной петли и вентральное ядро латеральной петли. К каждому из этих отделов идут проекции от определенных участков ядер первого уровня, а их эфференты адресованы среднемозговому акустическому центру. Кроме того, вентральное ядро латеральной петли дает начало нисходящим проекциям к моторным ядрам, осуществляющим контроль за голосовым аппаратом, т. е. опосредует видоспецифические вокализации. От верхней оливы начинается путь, обеспечивающий эфферентную иннервацию улитки. Особенностью, не получившей достаточных объяснений, является существование прямых восходящих путей к теленцефалону, источник которых ряд авторов обозначает как промежуточное ядро латеральной петли. Сложность идентификации комплекса латеральной петли у птиц, по-видимому, обусловлена его тесными связями с окружающей ретикулярной формацией, что не позволяет очертить границы некоторых участков конкретных ядер.**
В пределах класса млекопитающих имеются значительные экологически обусловленные адаптации слуховой системы и, соответственно, наблюдается разная степень ее развития. Центральные отделы слуховой системы оказываются весьма развитыми
95
уже в понтомедуллярном отделе, где располагается .комплекс кохлеарных (улитковых) ядер, подразделяемый у разных животных на два и более самостоятельных отдела. Весьма сложно организованы и верхнеоливарный комплекс, трапециевидное тело, комплекс латеральной петли, состоящие из нескольких ядер. От каждого из названных отделов берут начало восходящие проекции к среднему и промежуточному мозгу, многочисленные связи с ретикулярными отделами и моторными ядрами ствола, а также обратные связи в пределах сенсорного канала.
У однопроходных и сумчатых в акустической области продолговатого мозга описаны крупноклеточное и угловое ядра, nucll. magnocellularis et angularis. У однопроходных они подобны одноименным образованиям мозга рептилий как по характеру связей, так и по типам нейронов. По расположению же, в частности относительно веревчатых тел, они оказываются сходными и у сумчатых и не только с ядрами рептилий, но и птиц. У плацентарных эти структуры располагаются латерально относительно веревчатых тел и представлены передним вентральным кохлеарным ядром, nucl. cochlearis ventralis anterior, гомологичным крупноклеточному ядру рептилий, и дорсальным и задним вентральным кохлеарными ядрами, nucll. cochlearis dorsalis et ventralis posterior, гомологичными угловому ядру сумчатых. У млекопитающих происходит дальнейшая дифференцировка слуховых центров в области моста: усложняется комплекс верхней оливы, развиваются ядра трапециевидного тела, в системе латеральной петли выделяются дорсальное, промежуточное и вентральное ядра.* Все названные образования в мозге млекопитающих формируют многочисленные связи в пределах специфического сенсорного канала, а также проекции к другим отделам мезга (моторным ядрам черепно-мозговых нервов, ретикулярной формации, центральному серому веществу среднего мозга), обеспечивая тем самым организацию системных защитных реакций (в частности за счет активации мышц среднего уха) и видоспецифических вокализаций. От верхнеоливарного комплекса берут начало оливо-кохлеарные тракты, осуществляющие иннервацию рецепторных отделов.
Акустические центры млекопитающих отличаются от таковых у птиц и рептилий более разнообразным нейронным составом,
96
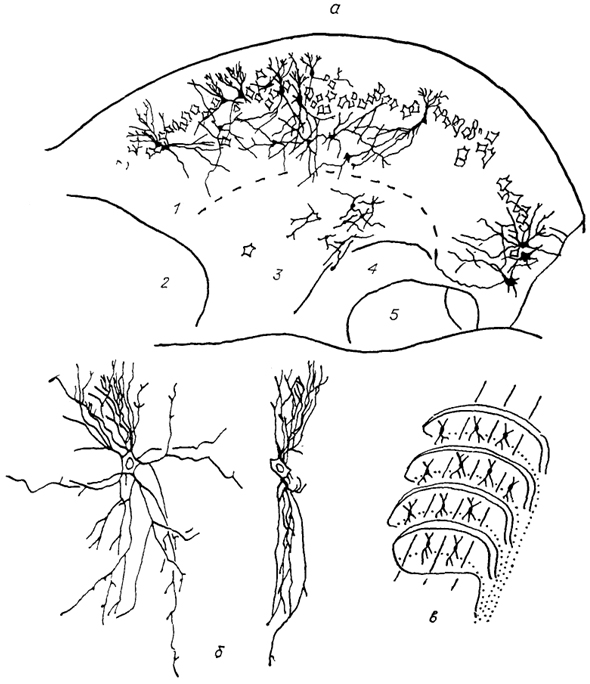
Рис. 40. Строение дорсального кохлеарного ядра кошки (Blackstadt e. а., 1984).
а - нейронный состав средней части дорсального кохлеарного ядра, б - пирамидоподобные нейроны, в - расположение пирамидоподобных клеток в ядре. 1 - слой гранулярных клеток, 2 - антеровентральное кохлеарное ядро, 3 - центральное ядро дорсального кохлеарного ядра, 4 - акустическая полоска, 5 - постеровентральное кохлеарное ядро.
многокомпонентностью и богатством связей внутри комплекса кохлеарных ядер и верхней оливы.
Среди кохлеарных ядер самым вариабельным оказывается дорсальное ядро. Оно наиболее развито у животных, ориентированных на восприятие звуков окружающей среды (хищные, зайцеобразные, грызуны), и имеет в этом случае четкое ламинарное строение (рис. 40). Напротив, в мозге эхолоцирующих млекопитающих, активность которых в значительной степени
97
обусловлена восприятием эха собственных сигналов, размеры дорсального ядра малы. Структурной особенностью этого ядра является наличие нейронов особой формы, напоминающих по ряду признаков клетки Пуркинье мозжечка. Они описаны под разными названиями (пирамидные, крупные биполяры, фузиформные) и располагаются в верхних слоях дорсального ядра перпендикулярно поверхности. Их дендриты ветвятся в одной, плоскости парасагиттально и пронизаны входящими под прямым углом афферентами. Аксоны этих клеток формируют слуховую полоску и проецируются на контрлатеральный задний холм. Другая особенность дорсального ядра - наличие разномодальных входов: помимо кохлеарных афферентов оно, в отличие от других улитковых ядер, получает проекции от главного ядра V нерва, вестибулярного комплекса и ядер дорсальных канатиков. Эфференты дорсального ядра направляются билатерально к верхнеоливарному комплексу, контрлатеральным ядрам латеральной петли и центральному ядру заднего холма. Ипсилатеральные проекции адресованы ретикулярной формации, медиальному и латеральному вестибулярным ядрам; и, вероятно, мозжечку.
Вентральные кохлеарные ядра проецируются на разные отделы верхнеоливарного комплекса. Кроме того, заднее вентральное ядро образует входы к моторному ядру V нерва ('иннервация небной занавески), двойному ядру (иннервация голосового аппарата) и ретикулярной формации. Эфференты вентрального ядра формируют трапециевидное тело, дорсального - слуховые полоски, corpus trapezoideum et striae acustica, соответственно. У некоторых млекопитающих показаны комиссуральные связи кохлеарного комплекса.
Верхнеоливарный комплекс млекопитающих значительно дифференцирован и имеет в качестве основных три отдела: медиальную верхнюю оливу, oliva superior medialis, латеральную верхнюю оливу, oliva superior lateralis, медиальное ядро трапециевидного тела, nucl. medialis corporis trapezoidei (рис. 41). У представителей разных отрядов степень их развития различна. Так, у приматов наблюдается постепенное увеличение размеров медиальной верхней оливы, у человека она достигает максимального развития. Латеральная верхняя олива и ядро трапециевидного тела велики у рукокрылых, насекомоядных, китообразных. У хищных и грызунов хорошо представлены все отделы комплекса.
Эфференты каждого из компонентов верхнеоливарного комплекса направляются к разным отделам вышележащих слуховых структур: ядрам латеральной петли и задним холмам. У некоторых видов описаны прямые проекции в таламус. Другие связи идут к моторным ядрам черепномозговых нервов (обеспечивая регуляцию вспомогательных структур периферического отдела слуховой системы), ретикулярной формации,
98
кохлеарным ядрам. Наконец, с этого уровня у млекопитающих осуществляется иннервация внутренних и наружных волосковых клеток волокнами ипси- и контрлатерального оливо-кохлеарных трактов соответственно.
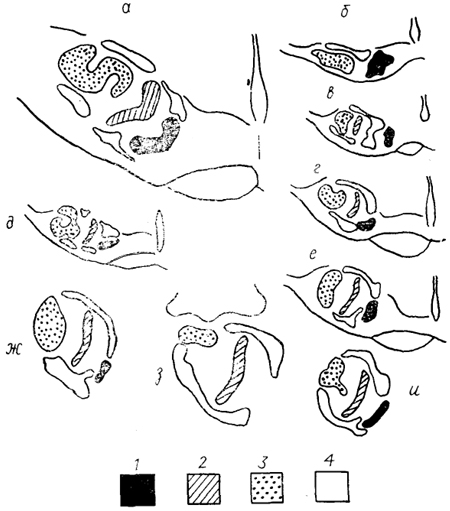
Рис. 41. Верхнеоливарный комплекс млекопитающих (Moore, 1987).
а - кошка, б - еж, в - землеройка, г - мартышка, д - галаго, е - совинолицая мартышка, ж - макака, з - человек, и - гиббон. 1 - трапециевидное тело; 2, 3 - медиальная (2) и латеральная (3) верхние оливы; 4 - периоливарные ядра.
Дорсальное ядро латеральной петли у млекопитающих характеризуется относительной стабильностью развития в мозге плацентарных. Оно получает входы от контрлатерального кохлеарного комплекса, ипсилатеральных медиальной верхней оливы и вентрального ядра латеральной петли, а также билатеральной верхней оливы. Эфференты дорсального ядра в основном восходят билатерально к задним холмам, образуя латеральную петлю, lemniscus lateralis, а также проецируются на контрлатеральный верхнеоливарный комплекс. Вентральное ядро латеральной петли слабо выражено у приматов и человека и сложно организовано у эхолоцирующих животных. Основные входы оно получает от кохлеарных ядер, а его эфференты в основном направлены к ретикулярной формации (что объясняет участие этой структуры в организации такой реакции, как startl-reflex), часть проекций восходит к задним холлмам.
99

Рис. 42. Схема преобразований акустико-латеральной системы позвоночных (Fritzsch, 1988).
1 - круглоротые (Lamvetra); 2-8 - рыбы: Polypterus (2), Squalus (3), Amta (4), Salmo (5), Latlmeria (6), Lepldoslren (7), Siren (8); 9-15 амфибии: Salamandra (9), Trlturus (10), Bolitoglossa (11), Ichthyophis (12), Xenopus (13), Rana (14), Bombina (15); 16 - рептилии (Testudo); 17 - птицы (Anas); 18 - млекопитающие (Didelphis).
У некоторых млекопитающих хорошо развито и промежуточное ядро латеральной петли, связанное с системой вокализации и участвующее в так называемом центральном ослаблении акустических сигналов у эхолоцирующих животных.
Отметим, что у всех амниот организация связей слуховой системы оказывается сходной. Так, центры первого и второго порядков (кохлеарные ядра, комплекс верхней оливы) формируют восходящий путь, адресованный среднему мозгу. Нейроны мезенцефалического уровня проецируются на дорсальный таламус, а через него на теленцефалои: в паллиальные (или стриатарные - у птиц) отделы. В процессе эволюции слуховой системы происходит усиление бинауральных взаимодействий, появление "спрямленных" восходящих и нисходящих путей.
Все ядерные формации слуховой системы амниот рассматриваются
100
как гомологичные, в то время как гомологизация их с подобной системой низших затруднена: убедительные доказательства единства происхождения слуховых центров мозга амниот и амфибий отсутствуют. Вопросы гомологизации не решены и в пределах группы рыб и между рыбами и амфибиями. Возможно, следует согласиться с авторами, допускающими независимость возникновения и параллельное развитие слуховых центров, по крайней мере, у амфибий и амниот. Что же касается рыб, то учитывая гетеромодальность рецепторов октаво-латеральной системы, трудно рассчитывать на обнаружение какой-либо предковой формы, общей для всех этих животных и давшей начало развитию акустической системы в разных группах (рис. 42).
Тем не менее не следует забывать, что между всеми названными системами - органов боковой линии, вестибулярной и слуховой - можно обнаружить значительное сходство. Оно касается и рецепторов, и путей проведения в мозге, и наличия специфических отделов, обеспечивающих эфферентный контроль рецепторных образований. Наконец, для акустической системы и системы органов боковой линии показано существование уровней, в которых обнаруживается полное представительство рецептивной поверхности и создаются "карты пространства" (в торусе и верхней оливе и в тектуме соответственно).
101
*
У низших позвоночных информация от соматических рецепторов передается не только от тройничного ганглия (по системе тройничного нерва), но и от сенсорных ганглиев VII, IX и X черепномозговых нервов. Однако у всех челюстноротых гипертрофия мандибулярной дуги отражается в доминировании сенсорного входа именно через V нерв (краниальный нерв мандибулярной дуги).
*
Комплекс этих черепномозговых нервов у низших позвоночных отчасти входит в состав соматосенсорной системы. Однако даже у рыб соматический компонент этих нервов иннервирует внешние покровы лишь в области наружных жаберных щелей. У четвероногих происходит дальнейшее уменьшение объема соматического компонента, а у млекопитающих единственной зоной, иннервация которой не обеспечивается системой тройничного нерва, является область наружного слухового прохода, иннервируемая клетками яремного ганглия.
*
Отметим, что волокна вестибуло-кохлеарного нерва адресованы и мозжечку.
**
Крупноклеточное вестибулярное ядро получает также входы нервов боковой линии.
*
Входящее в состав октаво-латеральной области амфибий дорсолатеральное ядро рассматривается как слуховое, и его описание будет дано ниже.
*
Для костистых рыб получающими акустическую информацию считаются тангенциальное ядро и ядро Дейтерса. У скатов Platyrhinoidis активность в ответ на предъявление звуковых стимулов зарегистрирована в ядрах октавного комплекса, и латеральной петле.
*
У некоторых рептилий обнаружены комиссуральные связи между ядрами кохлеарного комплекса.
*
Этим обстоятельством можно объяснить противоречивость сведений о комплексе латеральной петли птиц, включая утверждение о гомологии вентрального ядра птиц парабрахиальному комплексу грызунов, а дорсального - мезенцефалической ретикулярной формации.
*
При этом остается неясным вопрос об их гомологах в мозге птиц. Так, принято рассматривать медиальную верхнюю оливу как соответствующую ламинарному ядру, дорсальное ядро латеральной петли как гомолог вентральной части ядер латеральной петли птиц, а вентральное ядро латеральной петли млекопитающих - одноименному ядру птиц. Что же касается латеральной верхней оливы, медиального ядра трапециевидного тела и промежуточного ядра латеральной петли - ядер хорошо развитых у многих млекопитающих - то их гомологизация с мозгом птиц не проведена.
