Часть свободной энергии, высвобождаемой в окислительно-восстановительных реакциях, может быть использована для синтеза АТФ, который служит важнейшим, но не единственным переносчиком энергии в клетке. Помимо АТФ в метаболизме клетки участвуют другие богатые энергией соединения, к ним относятся другие нуклеотид ди- и трифосфаты, аденозинфосфосульфат, ацилфосфаты (например, карбамоилфосфат), фосфоенолпируват. Важнейшим макроэргическим соединением для анаэробов служит ацетил-КоА. Однако все эти соединения обмениваются с АТФ и поэтому представления об АТФ, как универсальном переносчике энергии, достаточно.
31
АТФ может синтезироваться путем субстратного фосфори-лирования, чаще всего за счет переноса фосфорильной группы от богатого энергией соединения на АДФ. Такой способ реализуется при гликолизе, разнообразных видах брожения и некоторых других процессах. Субстратное фосфорилирование осуществляется в цитоплазме и может быть воспроизведено в бесклеточных экстрактах.
Второй механизм синтеза АТФ, мембранное фосфорилирова-ние, необходимо зависит от наличия энергизованной мембраны и связан с использованием энергии трансмембранного электрохимического градиента ионов (в общем случае Δμl), чаще всего ионов водорода (Δμ,Н). Этот механизм реализуется в процессах дыхания (окислительное фосфорилирование) и фотосинтеза (фотофосфорилирование).
Окислительно-восстановительные реакции, в результате которых бактерии получают энергию, представляют собой перенос электронов от окисляемого вещества (донора электронов) к окислителю (акцептору). Перенос электронов от донора к акцептору осуществляется по градиенту редокс-потенциала через ряд последовательно функционирующих переносчиков. Часть из них закреплена в мембране, а часть находится в цитоплазме и сопрягает эти два компонента клетки. Набор переносчиков характерен для каждого организма.
Комплекс переносчиков электронов работает как универсальная структура для всех окисляемых веществ, лишь бы в метаболизме была реакция, совмещающая окислительное преобразование субстрата с цепью переноса электронов, на конце которой находится окислитель, например для О2 - цитохромоксидаза. Вместе с тем универсальным продуктом реакции должна быть генерация АТФ. Она осуществляется мембранным ферментом АТФ-синтазой, катализирующим синтез АТФ путем конверсии энергии трансмембранного электрохимического градиента протонов μH в энергию АТФ.
У аэробных органотрофов донором электронов служат органические вещества, окисление которых приводит к восстановлению переносчика водорода НАД в НАДН. Последний реагирует с электронтранспортной цепью. Моделью для окислительного фосфорилирования служит митохондрия эукариот (рис. 4).
32
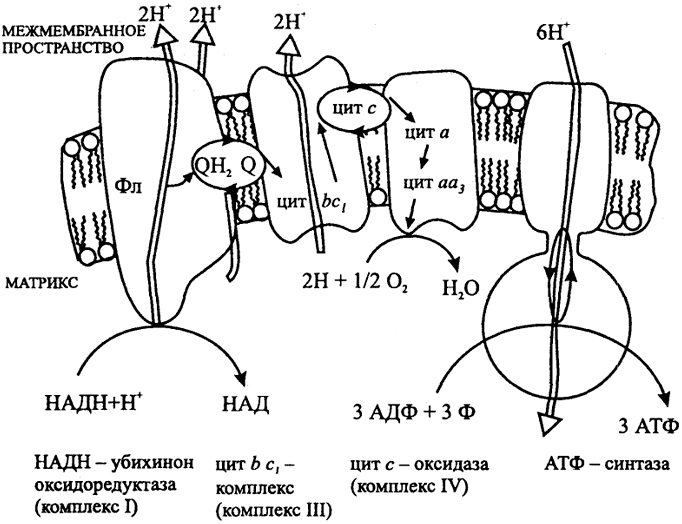
Рис. 4. Дыхательная цепь митохондрий
Цит - цитохром, Q - хинон, Фл - флавопротеин
Суммарный процесс переноса электронов разбит на три стадии, которые катализируются комплексами I, III и IV. Каждый комплекс представляет собой весьма крупное образование, построенное из многих полипептидных цепей. Первая стадия окисления НАДН катализируется комплексом I (НАДН: убихинон-оксидоредуктазой) и ведет к восстановлению убихинона Q. В состав комплекса I входит флавопротеин и FeS-кластеры. Вторая стадия, реакция окисления восстановленного убихинона и сопряженного восстановления цит с катализируется комплексом III (убихинон: цит с-оксидоредуктазой или вс1 - цитохромным комплексом). В его состав входит FeS-белок Риске, цит с1, или f, цит в. Третья стадия, перенос электронов от цит с на О2, катализируется комплексом IV, цитохромоксидазой. Перенос электронов сопряжен с переносом протонов через сопрягающую мембрану и генерацией ΔμН. Превращение энергии ΔμН в энергию АТФ осуществляется АТФ-синтазой.
У фотосинтезирующих организмов процессы преобразования световой энергии, приводящие к синтезу АТФ и образованию восстановителя, необходимого, например, для фиксации СO2, включают следующие этапы. Энергия света поглощается пигментами светособирающих антенных комплексов и передается в реакционный центр (РЦ), в котором происходит превращение электромагнитной формы энергии в энергию электрохимического
33
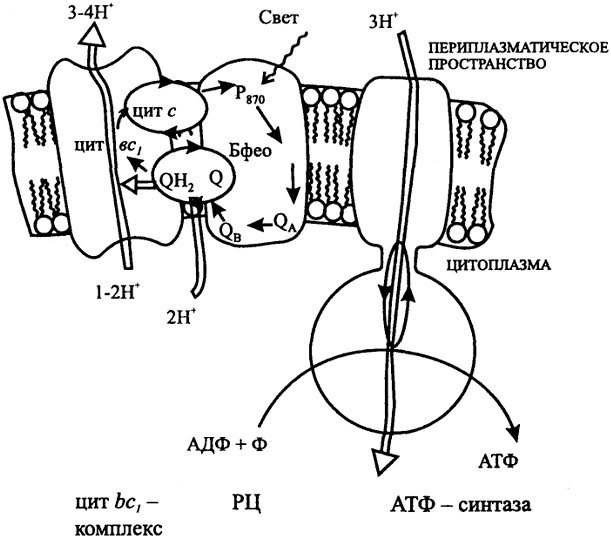
Рис. 5. Цепь переноса электронов при фотосинтезе А. Аноксигенный фотосинтез (пурпурные бактерии)
Р
870 - бактериохлорофилл; Бфео - бактериофеофитин; Q - хинон (Q, - плас-тохинон); цит - цитохром; Р
680' Р
700 - хлорофилл; Фео - феофитин; Фд - фер-редоксин; ПЦ - пластоцианин; ФС - фотосистема; РЦ - реакционный центр; ФНР - ферредоксин:НАДФ оксидоредуктаза.
Электроны от фотовозбужденного Р
870 передаются на убихинон Q, восстанавливая его, далее на цит bс
1, комплекс и через цит с и c
1, возвращаются к молекуле окисленного Р
870. Перенос электронов сопряжен с трансмембранным переносом протонов при окислении убихинопа цит bc
1, комплексом, что ведет к генерированию ΔμH. Результатом циклического переноса электронов является синтез АТФ.
разделения разрядов. Фотохимически активный хлорофилл (или бактериохлорофилл) РЦ, обозначаемый обычно буквой Р (от англ, pigment), переходит в электронно-возбужденное состояние и окисляется, отдавая электрон первичному акцептору. Перенос электрона на вторичный акцептор и восстановление окисленного Р первичным донором электронов стабилизирует разделение зарядов в РЦ. Дальнейшие процессы связаны с переносом электрона по электронтранспортной цепи, сопряженным с синтезом АТФ
34
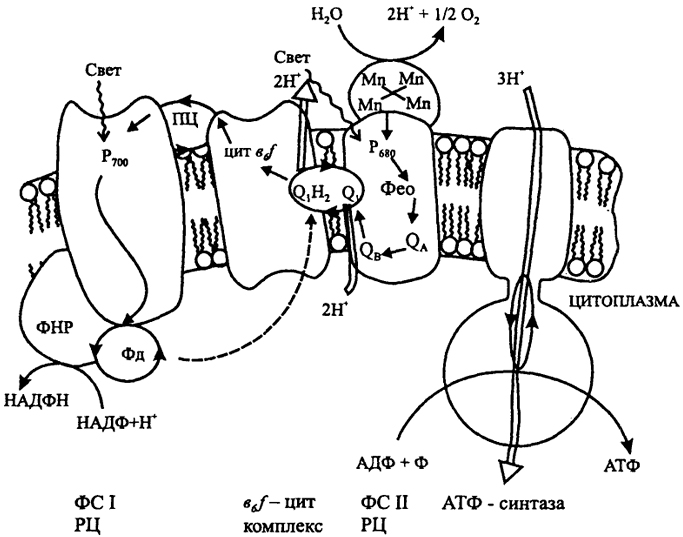
Б. Оксигенный фотосинтез
У оксигенных фототрофов имеются две фотосистемы: ФС I (с Р
700) и ФС II (с Р
680). В состав ФС II входит Mn-содержащий комплекс, осуществляющий разложение Н
2О. Электроны от фотовозбужденного Р
680 через ряд переносчиков РЦ передаются на пластохинон, далее в цит
b6f комплекс и через пластоцианин на Р
700. От фотовозбужденного Р700 (через ряд переносчиков РЦ) электроны поступают на Фд, связывающий ФС I с НАДФ-редуктазой (ФНР), которая восстанавливает НАДФ. Таким образом, происходит последовательный перенос электронов от воды к НАДФ. Реакции электронного транспорта, связанные с разложением воды в ФС II и окислением молекулы пластохинола цит
b6f комплексом, сопровождаются трансмембранным переносом протонов, сопряженным с генерированием ΔμH и синтезом АТФ. Образование АТФ возможно и в результате связанного с ФС I циклического преноса электронов (показан пунктиром).
(рис. 5 А, Б). Природа акцепторов электронов в РЦ и состав переносчиков электронов у разных групп фототрофов различаются. Существенно, что часть переносчиков являются общими для фотосинтетической и дыхательной электронтранспортной цепи, обусловливая у бактерий взаимосвязь процессов фотосинтеза и дыхания.
35
Различают циклический или нециклический пути переноса электронов. Циклический путь, свойственный, например, пурпурным бактериям, обеспечивает синтез АТФ (см. рис. 5 А), но не восстановителя. Поэтому восстановление НАД у этих микроорганизмов происходит в результате АТФ-зависимого обратного переноса электронов. У зеленых серобактерий перенос электронов осуществляется по нециклическому пути, обеспечивая синтез АТФ и восстановление НАД. Донорами электронов у аноксигенных, т.е. не выделяющих О2, организмов служат восстановленные соединения серы, S°, H2, Fe2+ или некоторые органические вещества. Цианобактерии и эукариотные фототрофы используют в качестве донора электронов Н2О и осуществляют фотосинтез с выделением О2 - оксигенный. В отличие от аноксигенных бактерий, имеющих одну фотосистему (ФС), у них последовательно функционируют две фотосистемы (см. рис. 5 Б) и может осуществляться как нециклический (так называемая Z-схема), так и индуцируемый ФС I циклический перенос электронов. Окисление Н2О осуществляется Mn-содержащим комплексом, связанным с ФС П. Особый тип фотосинтеза известен у экстремально галофильных архебактерий (галобактерий). В этом случае трансмембранный перенос Н+ и конверсия энергии света в энергию ΔμH с последующим синтезом АТФ осуществляются в результате светоиндуцированных преобразований бактериородопсина - пигмент-белкового комплекса, содержащего каротиноид ретиналь.
36
