Помимо межвидовой конкуренции за доминирование в пределах одной экологической ниши, существует конкуренция между группами организмов в пределах сообщества. Уже упоминалось, что предпочтение между акцепторами электрона для одних и тех же доноров определяется термодинамическими причинами с использованием в первую очередь реакций, дающих наибольший энергетический выигрыш. Наглядно этот тип конкуренции можно рассмотреть на уже неоднократно обсуждавшемся примере конкуренции между сульфидогенами и метаногенами, как это было сделано В.А. Вавилиным на основе данных микробиологов для очистных сооружений, где процесс идет в контролируемых условиях в очень большом масштабе.
Сульфидогены и метаногены имеют сходные доноры электрона - Н2 и ацетат, но разные акцепторы. При конкуренции следует учитывать кинетические свойства организмов. Если имеется рост, то используется кинетика Моно с Ks и μmax:
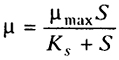
если же рост отсутствует, то кинетика Михаэлиса-Ментен с Кm и Vmax

232
В иле и в реакторах избыток сульфата приводит к полному подавлению водород-использующих метаногенов: нижнее пороговое значение Н2 для метаногенов 1,1 Ра, а для сульфатредукторов этот порог ниже и составляет 0,2 Ра.
Порядок использования Н2 и ацетата в анаэробном сообществе очень четкий: первым утилизируется Нз и только с большой задержкой начинает использоваться ацетат. Это не столь отчетливо видно по значениям μmах для чистых культур. В морских и пресноводных осадках ацетат потребляется в основном сульфатредукторами в условиях избытка сульфата. В метантенках это не так ясно. В морских осадках Desulfobacter postgatei имеет большее сродство к ацетату, чем Methanosarcina barken. Но в метантенках метаносарцины работают только в переходный период, уступая место метанотриксам, которые имеют большее сродство к ацетату. При оценке конкуренции следует учитывать и начальное соотношение биомассы конкурирующих организмов. Особенно важно это для медленно растущих организмов. Находящиеся в избытке организмы могут получить преимущество даже при худших для данных условий кинетических характеристиках.
Сульфатредукторы зависят от концентрации сульфата и скорости его поступления, что важно, например, в интерстициальной воде осадков. При высокой скорости потребления сульфата он может стать лимитирующим в биопленках с толщиной менее 1 мм. В отношении сульфата виды сульфатредуцирующих бактерий различаются: для морского Desulfobacter postgatei Km по сульфату составляет 200 мкМ, а у Desulfovibrio - 5-77 мкМ.
Конкуренция между использующими ЛЖК сульфатредукторами и синтрофными организмами сложна. При высокой концентрации сульфата эти сульфатредукторы растут быстрее синтрофов и являются основными потребителями ЛЖК. При низкой концентрации сульфата возможно разложение ЛЖК синтрофами на ацетат и водород с конкуренцией за эти субстраты между метаноге-нами и сульфатредукторами. Поскольку рост ацетогенов идет значительно лучше в кооперации с сульфатредукторами, создающими более низкий порог водорода, то вполне вероятен вариант, при котором гидрогенотрофные сульфатредукторы и ацетоклас-тические метаногены создают трофическую систему разложения
233
пропионата. В отсутствие сульфата сульфатредукторы могут функционировать как синтрофные ацетогены. В этом случае они растут с метаногенами как организмами, удаляющими водород.
Помимо сродства к субстрату, необходимо учитывать и влияние продуктов обмена. Токсичность сульфида определяется присутствием H2S, который может диффундировать в клетку; рH для диссоциации около 7, и поэтому небольшие вариации в рН, в пределах рН 6-8, могут оказывать сильное воздействие на сообщество. Сульфид подавляет рост метаногенов, но в гораздо большей концентрации, чем самих сульфидогенов. Так, свободный H2S подавляет метаногенез на 50% при концентрации 50-270 мг/л, а сульфатредуцирующих бактерий при 85 мг/л. Полное подавление Desulfovibrio происходит при 550 мг/л H2S. Выше рН 7,5 токсичность H2S резко снижается, вследствие доминирования ионов гидросульфида и сульфида. Однако в алкалофильном сообществе при рН 10,2 токсичность сульфида тоже проявляется очень резко и конечная концентрация сульфида в культурах алкалофильных сульфатредукторов обычно достигает не более 30 мМ. При этом не происходит угнетения продуцирующих Н2 спирохет. Если в сообществе присутствуют пурпурные бактерии, окисляющие H2S, то разложение органического вещества идет гораздо быстрее и концентрация H2S не поднимается выше лимитирующего уровня. В нейтрофильных условиях удаление H2S и снижение его токсического действия часто обусловлено связыванием H2S железом в нерастворимые сульфиды.
Другим примером конкуренции между функциональными группировками может служить конкуренция между гидрогенотрофными метаногенами и гомоцетатными бактериями. Организмы осуществляют реакции:
4Н2 + СО2 = СН4 + 2Н2О
4Н2 + 2СО2 = СН3СООН + 2Н2О
Метаногены, как уже упоминалось, обладают гораздо большим сродством к Н2, чем гомоацетатные бактерии, и поэтому в мезофильных условиях они выигрывают конкуренцию. В мезо-фильных условиях при температуре 18 °С основным продуктом
234
анаэробного разложения оказывается метан. Но при температуре ниже 15 °С высока скорость роста психроактивных гомоацетат-ных бактерий, и они становятся главенствующей группой, как было показано для анаэробного психрофильного сообщества тундры. В результате происходит переключение с метаногенеза на ацетогенез в зависимости от температурного порога, а не от набора субстратов. Таким образом, выигрыш зависит не только от сродства к субстрату, но и скорости роста при данных условиях, которая может быть резко различной; в приведенном примере - в зависимости от температуры. В условиях пониженных температур психроактивное сообщество переключается с метаногенеза на ацетогенез.
235
