Взаимодействие в трофических сетях между разнородными организмами обусловлено материальными потоками вещества между ними. Эти потоки зависят от скорости переноса вещества от организма А к организму В и, следовательно, от расстояния между ними. Перенос растворенных веществ между бактериями микронных размеров осуществляется путем молекулярной диффузии. Разобраться с процессами в пространственно-структурированном микробном сообществе удалось с помощью микроэлектродной техники, разработанной датскими учеными Б. Йор-генсеном (В. J0rgensen) и Н. Ревсбеком (N. Revsbeck) и широко примененной ими для природных сообществ. Ниже излагаются основные положения работы Йоргенсена.
217
Питание бактерий осуществляется исключительно путем молекулярной диффузии, движущей силой которой служит градиент концентрации вещества вокруг клетки. Он определяется соотношением:
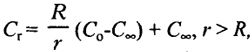
где Сr - концентрация на расстоянии г;
С0 - концентрация на поверхности клетки;
С∞ - концентрация в среде;
R - радиус клетки.
Максимальная скорость поступления вещества к клетке под действием молекулярной диффузии достигается при C0 = 0. Тогда общий диффузионный поток J к клетке:
J = 4πRDC∞ ,
где D - коэфициент диффузии, равный в воде для большинства мелких молекул примерно 10-5см2·с-1.
Объем клетки составляет - 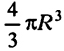 , и скорость метаболизма клетки, ограниченная скоростью поступления вещества к ней, составляет на единицу объема
, и скорость метаболизма клетки, ограниченная скоростью поступления вещества к ней, составляет на единицу объема . Отсюда поступление субстрата к клетке пропорционально
. Отсюда поступление субстрата к клетке пропорционально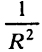 . Чем меньше размеры, тем эффективнее поступление субстрата. Бактерии размерного класса 1 мкм вообще не ограничены диффузией: за 1 с они извлекают вещества из объема, в 600 раз превышающего свой собственный.
. Чем меньше размеры, тем эффективнее поступление субстрата. Бактерии размерного класса 1 мкм вообще не ограничены диффузией: за 1 с они извлекают вещества из объема, в 600 раз превышающего свой собственный.
Уменьшение объема клетки, которое наблюдается у ряда морских, бактерий, обеспечивает более эффективное поступление вещества из крайне разбавленного раствора. В особенности это может быть важно для исключительно мелких фотосинтезирующих организмов "олиготрофного океана", лимитированных по азоту и фосфору. Они представлены недавно обнаруженным пикопланк-тоном очень мелких цианобактерий, относимых к синехококкам. Другим приспособлением служит изменение отношения "поверхность/объем" за счет образования выростов-простек или вытягивания
218
клеток в тонкие нити и увеличения пространства для расположения транспортных центров на поверхности клетки.
Расстояние L, которое проходит частица за время t, пропорционально корню квадратному из времени: L = √2Dt. Для бактерий размером в 1 мкм это время составляет 1 миллисекунду. Увеличение размеров влечет за собой транспортные ограничения. Время диффузии для молекулы составляет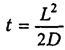 . Для молекулярной диффузии О2 в воде получены следующие значения:
. Для молекулярной диффузии О2 в воде получены следующие значения:
| Расстояние L |
1 мкм |
10 мкм |
100 мкм |
1 мм |
1 см |
10 см |
1 м |
| Время t |
0,3 мс |
30 мс |
30с |
4 мин |
7 час |
25 сут |
8 лет |
В коллоидах плазмы диффузия уменьшается в несколько раз. Очевидно, что в размерном классе более 10 мкм для перемещения вещества внутри клетки необходимы иные решения транспортных проблем, чем молекулярная диффузия. У эукариот, с характерными размерами клеток больше 10 мкм, задача решается локализацией процессов в структурах клетки, например митохондриях. В следующих размерных классах, помимо пространственной организации, необходимы специальные приспособления для транспорта. Нелинейный характер процесса приводит к появлению порогового значения размеров клетки, принимаемого как отличительный признак эукариот. Более крупные размерные классы организмов требуют решения транспортных проблем с участием тканей.
Перемещение клеток бактерий в жидкости с характерной скоростью 30 мкм/с не служит для облегчения поступления питательных веществ. Клетки останавливаются и меняют направление движения примерно каждую секунду. В этом случае их перемещение эквивалентно диффузии мелких молекул. Неподвижные бактерии в 1 мкм под действием броуновского движения имеют коэффициент диффузии 2·10-9см2·с-1, т.е. действительно неподвижны. Для подвижных бактерий коэффициент диффузии составит 10-5см2·с-1. Их движение с остановками и хаотическим изменением направления движения приводит к диффузии в
219
направлении субстрата, причем с более частыми остановками или увеличением скорости. Этот механизм создает микрозоны, где бактерии движутся особенно интенсивно и образуют тонкие слои, как это наблюдается, например, у Rhodospirillum. Перемещение становится выгодным для получения субстрата лишь у организмов размером около 40 мкм, т.е. крупного фитопланктона.
Турбулентность среды не влияет на молекулярную диффузию к бактериальной клетке, поскольку размеры вихрей составляют несколько миллиметров. Отсюда даже организмы размером в 100 мкм находятся в однородной среде.
Диффузионное поле вокруг частиц зависит от их размеров. Для бактерий размером 1 мкм понятие диффузионного пограничного слоя не имеет смысла, так как 10% от общего падения концентрации происходит на расстоянии 10 диаметров клетки. У агрегатов размером в миллиметры появляется уже гидродинамически обусловленный пограничный слой. Частицы "морского снега", размером примерно 3 мм, уже имеют пограничный слой, и снижение скорости движения воды на 10% происходит на расстоянии 9 радиусов частицы. Экспериментально показанный градиент кислорода падал в темноте от 280 мкМ в воде до 50 мкМ внутри частицы и возрастал на свету до 500 мкМ и выше за счет фотосинтеза диатомовых водорослей внутри частицы. Движение агрегата со скоростью 1 мм/с увеличивает его обмен со средой в 6-10 раз.
До сих пор мы рассматривали поведение взвешенных в объеме воды частиц, которые можно рассматривать как планктон. Иные отношения складываются для бентоса и для обрастаний, таких как биопленки, располагающихся на поверхности донных отложений. В толще воды господствует турбулентная диффузия Е, обусловленная движением вихрей, и выравнивающая концентрации в объеме. Примерно в 1 см от поверхности дна турбулентный перенос ограничивается вязкостью воды, и здесь господствует молекулярная диффузия. Примерно 1/10 от слоя, ограниченного вязкостью, составляет слой, получивший название диффузионного пограничного слоя 5е (diffusion boundary layer, DBL) и представляющего резкую граниду между областями турбулентной и
220
молекулярной диффузии. Толщина этого слоя 8е составляет примерно 0,5 мм, и уже на расстоянии в 1 мм от поверхности турбулентный перенос в 10 раз превышает молекулярную диффузию. Поверхность биопленки слизистая. В результате турбуленция Е вблизи поверхности биопленки затухает и на расстоянии долей миллиметра становится меньше молекулярной диффузии D. Над поверхностью образуется пограничный слой.
Движение воды над поверхностью приводит к утоныпению диффузионного слоя, который при больших скоростях может срываться. В общем, имеет место уменьшение толщины диффузионного слоя при повышении скорости течения над дном. Неровности рельефа приводят к изменению толщины диффузионного слоя, создавая большую мозаичность поверхности. Поэтому данные по пограничному слою относятся к таким поверхностям, как бактериальный мат, но не пористые песчаные осадки, при отсутствии тока грунтовых вод и, главное, отсутствии биотурбации донной фауной.
Диффузионный слой создает барьер между водой и осадком. При большой концентрации окисляемых веществ содержание О2 у поверхности осадка может быть низким даже при высоких скоростях потока воды, насыщенной кислородом. Такая ситуация наблюдается в областях осадконакопления, и тогда пурпурные бактерии создают слой у поверхности дна, а иногда на поверхность выступают и черные пятна сульфида. Кислород диффундирует через 0,2 мм пограничного слоя за 20 с, а через 1 мм за 480 с. Отсюда возникает задержка в переносе вещества диффундирующих через толщу осадка в воду и обратно. Перенос кислорода (260 мкМ) через диффузионный слой к поверхности, где он полностью поглощается микроорганизмами, имеет предел:
J =
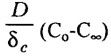
= 1 • 10
-10 моль • см
-2 • c
-1
или 94 ммоль·м-2·сутки-1 при экспериментально измеренной величине поглощения 10±30 ммоль*м-2сутки-1 для прибрежных осадков и 0,03±3? ммоль·м-2·сутки-1 для глубоководных осадков.
Увеличение скорости потока над биопленкой приводит к уменьшению пограничного слоя от более 1 мм в отсутствие движения
221
воды до 0,1 мм при скорости потока 8 см/с, обусловливая более резкий градиент, более глубокое проникновение О2 в мат. Это особенно важно для микроаэробов, как Beggiatoa и других градиентных организмов. Над неровной, поверхностью мата пограничный слой следует рельефу мата, но возникают микрозавихрения. Состояние пограничного слоя в масштабе 100 мкм удалось экспериментально измерить электрохимически с помощью микроэлектродов по ряду показателей: рН, рО2, H2S, N2O.
Внутри слизистой биопленки значения D, как оказалось, составляют только 50-60% от молекулярной диффузии. Для кислорода (Со - концентрация в среде) получается следующая картина:
- турбулентный поток =
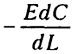 ;
;
- поток через пограничный слой =
 ;
;
- поток в осадке =
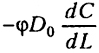 ;
;
- дыхание
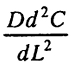
- Для стационарного состояния дыхание =

Отсюда по наклону кривой концентрации вычисляется дыхание. Градиенты могут быть очень резкими и зависят от концентрации веществ, в первую очередь от доступного органического вещества, используемого для дыхания. Например, в исследованном заливе у берегов Дании в период до оседания планктона кислород проникал на глубину 4 мм в осадок ила, а после "цветения" лишь на 1 мм. В цианобактериальных матах градиенты еще более резкие и составляют доли 1 мм.
Поскольку в сообществе происходит обмен продуктами и субстратами между организмами, то значение приобретает перенос вещества от одного организма к другому. Этот перенос осуществляется путем молекулярной диффузии. Перенос от точечного источника пропорционален квадрату расстояния. Поэтому для взаимодействующих организмов важно сближение на минимальное
222
диффузионное расстояние. На самом деле источник является не. единичным точечным, а некоторое число их распределено в трехмерном пространстве. Значит, оптимальным будет некое подобие кристаллической решетки. Для решения этой задачи биологические приспособления обусловили структурную организацию сообщества разнородных организмов, в котором действуют два противоположных требования. С одной стороны, размножение бактерий приводит к образованию микроколонии из однородных организмов, а с другой - необходимо взаимодействие различных видов.
Прямое микроскопическое наблюдение показывает, что в природных условиях образуются группировки разных организмов, часто объединенные общей слизью в микроагрегаты. Они особенно хорошо наблюдаются у водных организмов, начиная с первых шагов микроскопического исследования бактерий в природе в конце прошлого века Ф. Коном. Однако привычка микробиологов использовать для микроскопии препараты "раздавленной капли" сильно затрудняют анализ таких сложных агрегатов с характерными размерами в десятые доли миллиметра. Чтобы преодолеть это затруднение, Б.В. Перфильев разработал специальные приемы микроскопии с помощью плоскопараллельных капилляров. Это дало ему возможность иллюстрировать концепцию микрозонального развития микроорганизмов. Особенно наглядно микрозоны проявляются для железо- и марганецокисляющих организмов, окрашенных осадками окислов. Микрозоны создают следующую размерную ступень после требований, обусловленных минимальными диффузионными расстояниями в десятки микрон. Такое мозаичное распределение организмов в почве с помощью люминесцентной микроскопии продемонстрировал Д. Г. Звягинцев.
Примером реализации правила минимального диффузионного расстояния может служить организация метаногенного сообщества в очистных сооружениях. Для очистки жидких стоков сахароваренных заводов в Голландии в начале 1970-х гг. Г. Леттинга (G. Lettinga) спроектировал метантенки с восходящим потоком. Установка сначала работала немногим лучше обычных ферментеров с полным смешением. Но постепенно ее работа все
223
улучшалась и улучшалась, и наконец многократно превысила проектную эффективность. Оказалось, что ферментер заполнен гранулами миллиметрового размера. Когда гранулы этого осадка разрезали и посмотрели под микроскопом, обнаружили, что они представляют что-то вроде гаструлы и сложены рыхлой псевдопаренхимой метаносарцины. В межклеточном пространстве располагались разнообразные палочковидные бактерии, среди которых можно было опознать клостридии. В результате дальнейших исследований в гранулах было установлено наличие полного набора анаэробных организмов, необходимых для полного разложения различных органических веществ. В зрелом осадке (иле) матрицу гранулы представляли сплетенные в клубок нити мета-нотрикса, в петлях которого располагались другие организмы.
Другим примером подобных же отношений могут служить агрегаты бактерий в аэробной зоне, как хлопья псевдомонады Zoogloea ramigera в аэробных очистных сооружениях или нитчатых бактерий Haliscomenobacter во вспухающем иле аэротенков.
Наиболее сложную группировку составляет автономное сообщество цианобактериальных матов, где взаимодействуют многие группы разнообразных бактерий. В таких морфологически оформленных сообществах организмов большей частью можно выявить организм-эдификатор, морфология которого определяет структуру всего сообщества. Строительным компонентом для сообщества служит внеклеточная слизь, которая либо отстраняет другие организмы от микроколонии, например, от жгутов сплетенных трихомов цианобактерии Micfocoleus или жгутов серобактерии Thioploca, либо объединяет разные организмы в сообществе. Одним из вариантов объединения служит образуемая метаносарциной псевдопаренхима со свободными межклеточными пространствами. Иной тип создают нитчатые организмы, переплетение которых образует достаточно механически прочную структуру ткани со свободным пространством между нитями.
Резкие градиенты в сообществе создают оптимальные условия для транспортных процессов в межвидовом обмене метаболитами в сообществе. Вместе с тем микрозоны объясняют и возможность сосуществования, казалось бы, несовместимых процессов. Постоянной загадкой служит развитие анаэробных
224
организмов в зоне оксигенного фотосинтеза. Например, происходит образование метана в слое развития цианобактерий. Такое явление наблюдали в культуре цианобактерий из термофильного мата, подробно изучено выделение метана из клубков Trichodesmium в океане. Обычным является нахождение анаэробов в аэрируемом слое почвы. Наружный слой сферической гранулы (комочка почвы) сложен аэробными организмами. При достаточном поступлении растворимого субстрата из внутренней зоны, где разлагается нерастворимое органическое вещество, например растительные остатки, и вследствие медленной диффузии кислорода - в тысячу раз более медленной, чем в газовой фазе - внутри образуется анаэробная зона, где может развиваться анаэробное сообщество. Все это легко может быть подсчитано в модели и получило подтверждение микроэлектродными методами. Для нас важно, что размеры такой гранулы укладываются в миллиметровую шкалу.
Еще более наглядный пример представляют бентосные сообщества. В них последовательные этапы превращения веществ и последовательно действующие функциональные группы организмов располагаются послойно. В результате фронтального одностороннего поступления субстрата временная последовательность реакций превращается в пространственную. Трехмерная организация сообщества трансформируется в одномерную. Особенно тщательно организация сообщества была исследована на примере цианобактериальных матов.
Модель молекулярной диффузии является не единственной, хотя и наиболее важной. Кроме молекулярной диффузии, значение имеет конвекционный перенос вещества, вследствие, например, температурного перепада днем и ночью. Особые условия возникают, когда в среде имеется турбуленция и возможно перемешивание.
Гомогенное распределение клеток в объеме невыгодно для многих организмов, особенно для их взаимодействия в сообществе. Поэтому развиваются разнообразные приспособления для удержания на месте. Основным приспособлением служит гликокаликс - слизь или чехол. Он может быть важен для разных целей, например, для образования световых колодцев у
225
фотосинтезирующих организмов, для удерживания на расстоянии других организмов и образования диффузионных каналов. Важнейшим свойством гликокаликса является обобществление его для разнородных организмов и объединение их в морфологически оформленное сообщество. Образование таких сообществ является не досадным препятствием для выделения чистых культур, а биологическим приспособлением, заслуживающим специального изучения, которым оно стало лишь в последние годы. Помимо физической организации химических процессов в сообществе, морфология сообщества имеет своей целью максимальное удержание массы клеток без удаления из зоны реакции. Здесь микроорганизмы осуществляют "анти-хемостатную" стратегию.
В результате организмы развиваются не в водно-солевой среде, как принято думать, а в коллоидной матрице. Это обстоятельство часто не учитывается при биохимическом подходе. В биопленке создается закономерная морфологическая структура - "псевдоткань", сложенная разнообразными организмами. Приведенные примеры гранул в метантенках и, особенно, циано-бакте-риальных матов, иллюстрируют эту область существования сообществ организмов в природе.
226
