Низшие эукариоты относятся к протистам, преимущественно одноклеточным или колониальным микроскопическим организмам. Все они характеризуются наличием истинного ядра и компартментализации клетки. Некоторые протесты интересны как микроскопические хищники, способные захватывать твердые частицы и переваривать их внутри клетки. У них широко распространена способность к эндо-, эктосимбиозу и даже к слиянию с симбионтами в организм-химеру. Это сборная группа различного происхождения, разделяющаяся на много крупных таксонов - около 40 классов - с не очень четкой номенклатурой, варьирующей у разных авторов.
Происхождение протистов основывается на сопоставлении "16S рРНК-подобных молекул", причем они оказываются очень разнообразными. Дерево протистов предложено М. Согиным (М. Sogin) в 1991 г., и в нем лишенные митохондрий организмы оказались у основания дерева. Отсюда делается предположение, что митохондрии представляют позднейшее приобретение у первоначально анаэробных протистов. Включение бактериального эндосимбионта - предшественника митохондрий произошло до
176
появления эвгленид, представляющих важнейший шаг в эволюции протистов. Euglenozoa включают гетеротрофные Kinetoplastida (включая свободноживущие Bodonida и паразитические Тгурапо-somatida), причем автотрофные эвглены в качестве фотосинтезирующего симбионта имеют не цианобактерии, а зеленые эукариотные водоросли, т.е. их симбиоз представляет позднее событие. Миксомицеты и акразии появляются на дереве еще позднее. Паразитические формы у них утратили митохондрии вторично.
Протесты разделяют на Algae и Protozoa по наличию или отсутствию хлоропласта и соответственно фототрофному ("растительному") или органотрофному ("животному") способам питания. Эта функциональная система более не соответствует представлению о протистах. Филогенетическая схема разделения их на группы изображена на рис. 30. Здесь представлена попытка построить "лестницу существ" для протистов по мере усложнения. На самом деле филогенетическая система протистов, более чем любой другой группы, представляет комбинаторную сетку, вследствие самого способа возникновения низших эукариот как химер. В рассмотрении этих групп мы сосредоточим внимание на свободноживущих организмах, представляющих приоритет для природоведческого подхода.
Дипломонады. Первоначальными считают организмы, лишенные митохондрий, а сейчас это анаэробы пищеварительного тракта, как лямблии (Giardia intestinalis). Интереснее симбионты термитов, способные захватывать кусочки древесины и анаэробно сбраживать ее до ацетата, который и усваивается насекомым. Эти организмы очень склонны к симбиотическим ассоциациям с бактериями, например, с покрывающими их поверхность спирохетами, которые функционируют как их органы движения.
Остается неясным, почему предполагаемые первичные анаэробные протисты сохранились лишь как паразиты и симбионты, и почему исчезли из анаэробных мест обитания, где большинство анаэробных протистов развивается как вторичные вселенцы за счет симбиоза с прокариотами.
Кинетопластиды и эвглены. У основания дерева свободно-живущих протистов, обладающих митохондриями с дисковидными кристами, и соответственно аэробных, располагаются эвглеиовые.
177
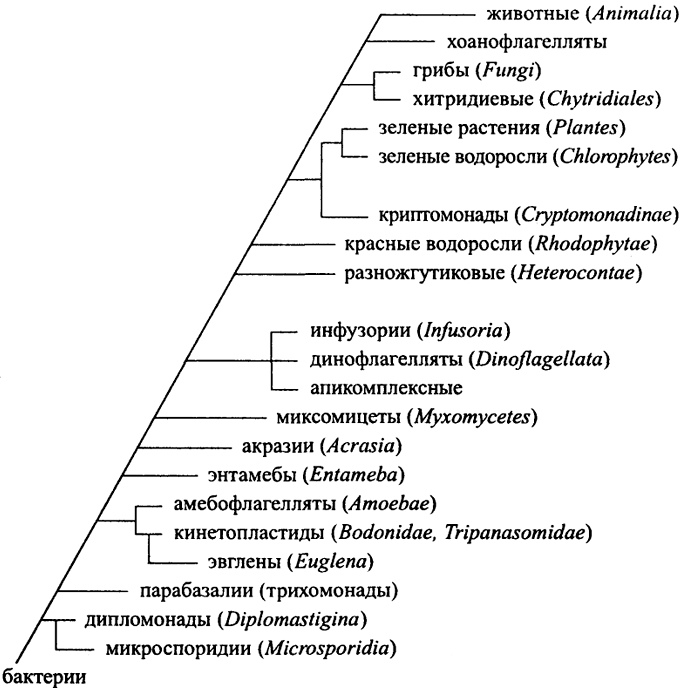
Рис. 30. Вариант филогенетического дерева эукариот на основании сравнения последовательностей рибосомальных РНК (По J. Perry, J.T. Staley. 1997)
Их питание очень разнообразно: фототрофное, фаготрофное, сапротрофное. Фототрофное питание осуществляется за счет захваченной и инкорпорированной зеленой водоросли с хлорофиллами а и b, причем она довольно легко теряется. Фаготрофные эвглены захватывают добычу с помощью микротрубочек глотки, расположенной рядом с резервуаром, откуда выходит жгутик. Наконец, сапротрофное питание осуществляется за счет развития в местах, обильных органическим веществом - мезосапробной зоне.
178
К эвгленам близки кинетопластиды, имеющие своеобразную организацию митохондриальной ДНК в виде кинетопласта - сетки связанных между собой кольцевых структур. Несмотря на необычность организации генома, эти протисты процветают и представлены широко распространенными в загрязненных водах и почве бодонидами. Bodo имеет глотку из микротрубочек и питается бактериями. Паразитическими родственниками его являются Leishmania, Trypanosoma - возбудители тяжелых заболеваний.
Третью ветвь в этой группе составляют распространенные в почве, пресной воде и море амебофлагелляты. При снижении уровня питательных веществ амебы переходят в жгутиковую стадию. В отличие от истинных амеб, они вытягивают единственную псевдоподию. Большинство из них питается бактериями. Размножаются они очень быстро.
Следующими на филогенетическом дереве появляются миксомицеты. Их объединяют по трубчатой форме митохондриаль-ных крист. К грибам они никакого отношения не имеют и название возникло по внешнему сходству их плодовых тел. Миксомицеты - это почвенные амебы, плодовые тела которых обеспечивают стадию расселения спор токами с воздуха аналогично тому, как это происходит у настоящих грибов. Собственно миксомицеты образуют плазмодий. Он образуется слиянием гаплоидных жгутиковых в диплоидную стадию, разрастающуюся в огромную многоядерную амебу - плазмодий.
Акразии, как Dictyostelium, напоминают миксомицеты, но остаются в виде отдельных одноядерных амеб, обладающих, однако, способностью к кооперативному действию при образовании плодового тела.
Объединение следующей группы протистов основано на присутствии под унитарной мембраной альвеол - слоя мелких пузырьков; они особенно выражены у инфузорий, а у динофлагел-лят образуют целлюлозные пластинки клеточной стенки под мембраной.
Динофлагелляты представляют важнейшую и разнообразнейшую группу протистов, преимущественно фототрофных. Помимо свободноживущих планктонных форм, они дали начало
179
симбиотическим "зооксантеллам", превратившим ряд других протист, как фораминиферы, и низших многоклеточных организмов, как кишечнополостные и губки, в фотоавтотрофные симбиотические консорции. Морской фитопланктон в значительной степени представлен динофлагеллятами.
Инфузории стали наиболее известной линией крайнего морфологического усложнения одноклеточного организма. Классификация их основывается на морфологии, хотя вследствие крупных размеров относительно несложно получить их монокультуры и даже клоны. Для очистки от бактерий широко применяются антибиотики. Инфузории в основном хищники, питающиеся бактериями или другими протестами. Первая чистая культура инфузорий была получена 21-летним А. Львовым (A. Lwoff) в 1923 г., и это была Tetrahymena, послужившая затем моделью для исследований протист, аналогично чистым культурам бактерий.
Следующие группы организмов представляют предмет альгологии, и только их наиболее простые одноклеточные представители попадают в группу протистов-микроорганизмов.
Разножгутиковые составляют важнейшую группу фототро-фов, доминирующую в море и столь же значимую для него, как растения для суши. К ним принадлежат золотистые водоросли, например Ochromonas, ксантофиты от Botrydiopsis до Vaucheria, отвечающие примерно за 20% первичной продукции, диатомовые, многоклеточные бурые водоросли-макрофиты. Разножгутиковые интересны тем, что составляют ряд усложняющихся морфологически форм от одноклеточных жгутиковых до гигантских морских водорослей со сложным дифференцированным талломом. Хлоропласты их имеют пачки мембран, собранные по три, и их наружная мембрана связана с эндоплазматическим ретикулумом и через него с ядерной мембраной.
Из бесцветных разножгутиковых большую группу составляют водные "грибы" - оомицетьг, например Saprolegnia, по своим молекулярным характеристикам не имеющие ничего общего с собственно грибами.
Криптомонады представляют фототрофных одноклеточных жгутиковых, фотосинтетический аппарат которых, скорее всего, представляет дегенерировавшую эукариотную водоросль. Эта
180
малая группа рассматривается сейчас как химера - продукт симбиогенеза.
Красные водоросли образуют такой же параллельный ряд усложняющихся форм, как бурые водоросли, но не имеют жгутиковых стадий, а их пигментный состав отличается от других окси-генных фототрофов и ближе всего к цианобактериям. Их хлоропласты сложены отдельно располагающимися мембранами с фикобилисомами на них, как у цианобактерий. Это преимущественно морские бентосные организмы.
Зеленые водоросли составляют наиболее разнообразную группу "классических" оксигенных фототрофных организмов. Они доминируют и на суше, и в континентальных водах. Последовательный ряд морфологического усложнения при одинаковом типе обмена прослеживается здесь с наибольшей очевидностью. Микроводоросли, такие как Chlorella, со времен Бейеринка стали объектом микробиологических исследований в чистой культуре. Хлоропласты их имеют пачки мембран и включают пиреноид - место синтеза крахмала. Зеленые водоросли достигают крупных размеров и сложной дифференциации у харовых водорослей, представляющих вероятных предшественников наземных растений.
Наземные растения - Plantae - включают как сосудистые растения, так и криптогамовые. Они появляются на Земле относительно поздно - в силуре - в условиях уже развитой биосферы со сложившимися биогеохимическими циклами и осуществляющими разнообразные реакции организмами. Отличительной особенностью растений, в отличие от водорослей, является вынесение фотосинтезирующего аппарата в виде листьев в атмосферу к свету, что потребовало создания устойчивого к разложению органического опорного "скелета" на основе лигноцеллюлозы. Растения становятся обитателями аэротопа - прилегающего к поверхности Земли слоя атмосферы, пронизанного стеблями растений.
Корень дерева животных - Animalia - связывается с одноклеточными и колониальными хоанофлагеллятами. Это мелкие преимущественно морские протисты-фильтраторы. Они имеют венчик липких щупалец, к которым прилипают частицы при биении единственного жгутика, создающего ток воды. Клетки
181
хоанофлагеллят иногда окружены корзинкой-лорикой. Точно такие же клетки составляют основу пищевого аппарата губок. От колониальных эукариот начинается хорошо разработанная зоологами систематика беспозвоночных животных, основанная на наличии у них для питания твердыми частицами пищеварительного тракта с населяющими его микроорганизмами.
Следующую группу эукариот составляют грибы - объект микологии. Их ближайшие родственники - связывающие их с протистами хитридиомицеты. Это водные организмы со жгутиковыми зооспорами. Тело их представлено либо одной клеткой, либо неправильно ветвящейся массой, напоминающей мицелий.
Истинные грибы, Fungi - наземные организмы, мицелий которых сходен с мицелием прокариотных стрептомицетов и представлен длинными ветвящимися гифами - трубками со сложенной хитином оболочкой. Мицелий составляет один большой организм. Несмотря на то, что грибы относят к микроорганизмам, они на самом деле представляют, формально говоря, самые крупные организмы на Земле. Мицелий грибов приспособлен для развития в почве и способен переносить контакт с воздухом, нередко образуя воздушный мицелий; поэтому грибы выигрывают у бактерий при переходе к засушливым (ксерофитным) условиям. Грибы размножаются неподвижными спорами, разносимыми ветром, и соответственно способу размножения разделяются на аскомицеты и базидиомицеты. Сеть мицелия обеспечивает возможность транспорта по ней питательных веществ от клетки к клетке. Грибы - строго осмотрофные организмы с жесткой клеточной оболочкой, не допускающей возможности проникновения твердых частиц. Вместе с тем грибы обладают мощным аппаратом гидролитических экзоферментов, обеспечивающих им возможность деградации полисахаридов растительных оболочек (и, что особенно важно, лигнина), липидов, нерастворимых азотистых соединений. Они составляют группу сапрофитных микроми-цетов, особенно богатых в лесной подстилке. Многие грибы вступают в симбиотические и паразитические отношения с сосудистыми растениями, так что их можно рассматривать как постоянные компоненты растительно-грибной системы. Так называемые лихенизированные грибы, имеющие морфологически
182
оформленные симбиотические группировки с микроводорослями, получили название лишайников. С грибами как важнейшими деструкторами в углеродном цикле связано функционирование наземных экосистем. Некоторые грибы стали лабораторным объектом микробиологии и биотехнологии, как хорошо известный продуцент пенициллина Penicillium. Сейчас известно несколько сот тысяч видов грибов и, конечно, такая обширная и важная группа служит объектом специальной дисциплины - микологии.
Из приведенного очень беглого обзора низших эукариот с очевидностью следует, что их морфологическое разнообразие подчиняется общим законам дифференциации в большей степени, чем общности происхождения. Филогенетическая система на основе последовательностей РНК, приведенная здесь, отвлекает от независимых от происхождения законов морфологической дифференциации. Функциональные особенности протистов в гораздо большей степени, чем у бактерий, зависят от их морфологии. Типы питания протист определяются дыханием митохондрии и/или фотоавтотрофией хлоропласта. Между тем все остальные особенности, позволяющие им занять разнообразные ниши, связаны с физическими факторами. Растения, грибы и животные представляют продукты усложнения очень немногих исходных групп протистов, и их филогению можно представить в виде деревьев, возвышающихся над кустарником многочисленных групп протистдв. Для многоклеточных эволюция связана с последовательной дифференциацией на основе морфологического усложнения в монофилетических линиях и явно связана с увеличением размеров, требующим нового способа решения проблем транспорта внутри организма, обеспечиваемого сосудистой системой растений и подвижностью пищи у животных, например, в пищеварительном тракте. Характерной особенностью протистов, в отличие от происходящих от них многоклеточных организмов, оказывается способность к многократному последовательному усложнению путем комбинирования в разных типах симбиогенеза. Они принципиально полифилетичны.
Геохимической особенностью протистов стала отсутствующая у прокариот способность к построению минерального скелета
183
с участием внутриклеточных органелл. В результате в биогеохимические циклы оказались вовлечены такие элементы, как кальций, кремний, стронций, которые у прокариот участвовали в циклах лишь через биологически опосредованные реакции.
В функционирование биосферы протесты привнесли способность питаться твердыми частицами. В роли твердых частиц оказались бактерии, составлявшие, вероятно, первоначальную пищу эукариот-протистов. Поскольку это принципиально иной тип питания, чем фотосинтез зеленых растений, гидролитическая активность грибов и даже всасывание пищи в результате ее переваривания в пищеварительном тракте, его следует рассмотреть специально как фаготрофию - способность осуществлять внутриклеточное переваривание.
184
