По отношению к солености микроорганизмы разделяются на несколько групп (рис. 28). Соленость действует на клетки как осмотический фактор. Для своего существования клетки должны поддерживать тургор: мембрана должна плотно прилегать к клеточной стенке, как пластиковый мешок с жидкостью к сетке. При нарушении этого состояния происходит плазмолиз. Организмы разделяются на физиологические группы в соответствии с осмотическими условиями среды.
Считается, что обитатели пресных вод чувствительны к 3,5%-ной концентрации NaCl, как в морской воде, и в описание организмов обычно входит такой тест. Этот тест не совсем справедлив в отношении обитателей почвы, где концентрация почвенного раствора резко меняется в зависимости от дождей или засухи. Почвенные организмы должны приспосабливаться к резким изменениям осмотических свойств среды обитания. Многие из них являются галотолерантными. Наиболее характерны здесь грамположительные организмы. Обитатели ультрапресных вод развиваются в среде с содержанием солей ниже 100 мг/л, в том
114
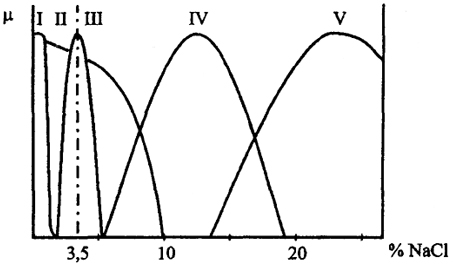
Рис. 28. Отношение к солености роста разных групп микроорганизмов
I - ультрапресные; II - галотолерантные; III - морские; IV - умеренные гало-филы; V - экстремальные галофилы; μ - скорость роста
числе в дистиллированной, дождевой воде или в воде сфагновых болот. Примером могут служить Caulobacter, Spirillum.
Морские организмы развиваются при солености, равной 2-4%, причем под соленостью здесь понимается именно содержание NaCl. Обозначение морских организмов как галофилов не совсем удачно, так как они не приспособлены к значительному повышению солености. Среди морских организмов обильны про-теобактерии, например псевдомонада Alteromonas.
Галофилы развиваются при солености, существенно превышающей соленость морской воды. К умеренным галофилам относятся организмы, имеющие верхний предел солености до 15%.
Экстремальные галофилы развиваются вплоть до насыщения воды NaCl при 30%-ной солености. К ним относятся архебак-терии сем. Halobacteriaceae и некоторые галоанаэробы. Наиболее обильное развитие галофилов происходит при солености 10-20% в испаряющихся лагунах. Здесь их разнообразие очень велико, включая цианобактерии, прежде всего Microcoleus, ряд про геобактерий, как Halomonas, специальную группу Haloana-erobiales. Соленость выше 10-15% исключает развитие эукариот, кроме немногих вторичных вселенцев, как зеленая водоросль Dunaliella.
От морских галофилов следует отличать обитателей высокоминерализованных континентальных вод, как правило, с повышенным, иногда вплоть до насыщения (примерно 25%) содержанием
115
соды. Поскольку такие воды обычно имеют и высокое значение рН и содержат NaCl, обитателей их относят к галоалкало-филам, учитывая, что они приспособлены и к высокой минерализации и к рН. По сути, обе группы следовало бы отнести к "натрофилам", поскольку они нуждаются в высокой концентрации Na+.
Организмы, развивающиеся при высокой концентрации органических веществ, обычно называют осмофилами, в большинстве это обитатели сиропов. Среди них много эукариотных организмов, таких как мицелиальные грибы и дрожжи.
Все прокариоты, за исключением воздушного мицелия акти-номицетов, развиваются в водной среде. Испарение воды определяется водным потенциалом aw, представляющим отношение давления водяного пара над раствором по отношению к давлению над дистиллированной водой. Соответственно обитатели ультрапресных вод развиваются при потенциале 1, для морской воды этот потенциал равен 0,98, для гиперсоленых озер - 0,7. Понижение водного потенциала за счет засолки, засахаривания, сушки служит способом сохранения пищевых продуктов от развития обычных организмов.
Для почвенных организмов значение имеет ксерофилия, т.е. приспобление к сухости. Более приспособленными оказываются грамположительные организмы, в особенности актиномицеты. Наиболее приспособлены к сухости грибы, мицелий которых выходит в воздушную среду, например у лишайников. Бактерии переносят сухость, создавая гидрофильные капсулы вокруг колоний; типичный пример такой стратегии представляют почвенные цианобактерии, например Nostoc. Другим способом перенесения сухости служит образование спор, обладающих дополнительным свойством термостабильности, конидий у актиномицетов, разного рода цист, служащих не только для переживания, но и для расселения. Организмы в переживающих стадиях способны широко распространяться по воздуху с пылью, обеспечивая повсеместность присутствия бактерий на земном шаре.
Приспособление к осмотическому состоянию среды осуществляется с помощью синтеза низкомолекулярных органических веществ, относительно устойчивых метаболически в цитозоле, пассивно удерживающихся мембраной. Такие вещества получили
116
название осмопротекторов или осмолитов. К ним относится широкая категория веществ, различных у разных групп организмов. Из аминокислот такую роль играют пролин и глутамат, из Сахаров - трегалоза, свойственные слабым галофилам. Наиболее распространенным осмопротектором служит бетаин, свойственный и почвенным, и умеренно галофильным морским организмам. Для морских водорослей важным осмопротектором служит метилсульфопропионат. Галофильные цианобактерии синтезируют гликозил-глицериды. Важным и характерным осмопротектором служит иминокислота эктоин. Во всех случаях для поддержания осмотического равновесия организмы вынуждены синтезировать возрастающие количества осмопротекторов, составляющие существенную часть биомассы. Эти осмолиты называют совместимыми, потому что они не нарушают энзиматической активности в клетке и даже служат протекторами от других неблагоприятных влияний. Иная стратегия наблюдается у архебактерий Halobacte-riaceae и эубактерий Haloanaerobiales, где в клетке накапливается ион К+, и их специфические ферменты работают в солевом растворе. У эукариот роль осмопротекторов выполняют полиспирты, такие как сорбит, маннит, свойственные лишайникам, или как глицерин у Dunaliella. Очевидно, что синтез осмопротекторов представляет характерную адаптивную реакцию.
117
