Размножение делением надвое описывается простой формулой
Nt = N0·2n,
где N, - конечная;
N0 - начальная численность биомассы;
n - число удвоений за время t.
| Отсюда n = |
lg Nt - lg N0 |
|
| lg 2 |
|
или приближенно n = (lg Nt · lg N0) · 3,32
Время удвоения или генерации составляет t/n.
74
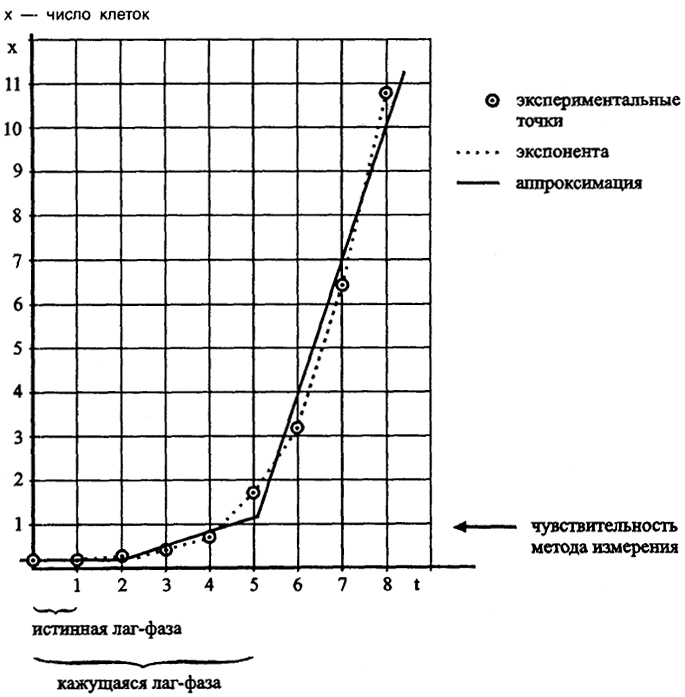
Рис. 22. Аппроксимация экспоненциального роста прямыми
Если изобразить этот процесс на графике, то получается кривая экспоненциального роста, которая в полулогарифмических координатах lg N/t превращается в прямую. На практике экспоненциальный рост может быть аппроксимирован двумя прямыми по трем точкам (рис. 22) и воспринимается как пороговое событие с внезапным переходом в состояние насыщения по достижении максимальной плотности. Начальная фаза экспоненты обычно воспринимается как лаг-фаза, отчасти в связи с порогом распознавания начала роста по мутности при lg Nt = 5 или 6. От этой кажущейся лаг-фазы следует отличать истинную лаг-фазу, когда размножения нет и происходит приготовление клетки к началу
75
размножения инициационными процессами внутри клетки. Лаг-фаза определяется отрезком времени, отсекаемым на полулогарифмическом графике прямой численности живых клеток. По мере приближения к предельной численности рост замедляется. Предельная численность бактерий микронных размеров достигает 1012 клеток в см3; вес сырой массы одной бактерии составляет n·10-13 г. К таким плотностям приближается масса бактерий в колониях и биопленках, но не в жидких культурах. Обычно максимальная численность определяется использованием питательного субстрата, но могут быть и иные причины, например отравление продуктами обмена. Есть и более тонкие механизмы регуляции плотности популяции с использованием сигнальных молекул, повышение концентрации которых в среде воспринимается генетическим аппаратом бактерий. По достижении максимально возможной в данных условиях плотности, бактерии переходят в стационарную фазу, характеризуемую иным физиологическим состоянием, обычно обозначаемым как анабиотическое. При этом в клетке резко снижается содержание рибосом и соответственно РНК. Под микроскопом в фазовом контрасте такие клетки выглядят бледнее и они иначе окрашиваются флуорохромами. В стационарной фазе только часть клеток сохраняет способность к размножению. При пересеве в свежую среду неразмножающиеся клетки создают начальный фон, маскирующий начало экспоненциальной фазы, а перестройка клеток вызывает лаг-фазу. Вслед за стационарной фазой наступает фаза отмирания, сопровождающаяся или не сопровождающаяся лизисом. Отмирание бактерий - процесс, связанный с метаболической активностью. Если эта активность заторможена, например, низкой температурой, то клетки могут сохранять жизнеспособность очень долго, например, в вечной мерзлоте или при криоконсервации. Сохранение жизнеспособности оценивается сроком заведомо больше десятков тысяч лет во льдах Антарктиды и более миллиона лет для вечной мерзлоты. Таким образом, отмирание - активный процесс, и обычно его описывают как постоянную гибель определенной части клеток за единицу времени, что дает на полулогарифмическом графике нисходящую прямую. Отмирание бактерий в естественных условиях приводит к так называемому самоочищению природных
76
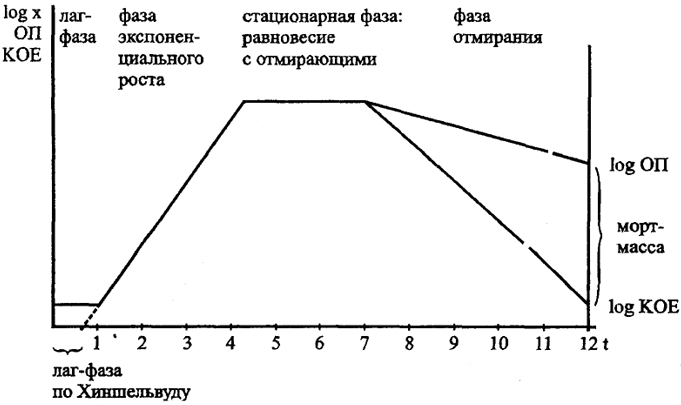
Рис.23. Кривая роста бактерий
ОП - оптическая плотность культуры, КОЕ - число колониеобразующих единиц, х - число клеток
сред. Процесс отмирания не менее важен для поведения организма в природе, чем рост, но изучен гораздо менее подробно. Внимание привлекли уже упоминавшиеся "некультивируемые формы", которыми называют организмы в состоянии неготовности к прорастанию на средах, обычно пригодных для их роста.
Теоретически кривая роста бактерий на полулогарифмическом графике (рис. 23) описывается трапецией, сдвинутой от начала координат на величину лаг-фазы. Время достижения максимальной численности зависит от размеров инокулята, причем здесь дело не только в арифметических расчетах, но и в биологических особенностях, когда при достаточной начальной плотности быстрее проходит лаг-фаза.
Между синтезированной биомассой и количеством потребленного вещества (или образованных продуктов) существует пропорциональность, обозначаемая как "константа урожая" Y. Для аэробных органотрофов эта величина составляет около 50 весовых процентов. Обычно Y оценивается в граммах сухой биомассы на 1 моль использованного субстрата. Очевидно, что эта величина зависит от термодинамического выхода основной реакции
77
катаболизма. Более точная величина получается для YАТP - количества синтезированной биомассы (в граммах) на 1 моль АТФ, образованного в процессе метаболизма, - равного приблизительно 10 г. Это универсальная оценочная величина для экспоненциального роста, хотя у некоторых организмов она может заметно отличаться. Пропорциональность между ростом и потреблением субстрата и стехиометрическим образованием продукта позволяет оценивать рост химическими методами там, где подсчет клеток затруднителен, например, для нитрификаторов первой фазы по образованию, а второй - по потреблению нитрита. В стационарной фазе роста идет потребление субстрата без размножения, и затраты на этот процесс обозначаются" как "энергия поддержания". По мере увеличения плотности бактерий она составляет все большую часть баланса. Очевидно, что при активном росте концентрация субстрата в среде снижается до минимума, и поэтому нельзя ожидать высокой концентрации его там, где выросли бактерии.
78
