Генетическая информация записана в виде пар нуклеотидов на двойной спирали ДНК. Блок информации, соответствующий какому-либо свойству или признаку, обозначают как ген. Полный набор генов называется геномом. Геном бактерий включает несколько тысяч генов, расположенных линейно на макромолекуле ДНК, называемой хромосомой. У прокариот имеется одна кольцевая хромосома на клетку (за исключением организмов, у которых не произошло деление и образовались, например, нитчатые формы), не отделенная от цитоплазмы мембраной и видимая на ультратонких срезах в виде клубка нитей в светлом пространстве (нуклеоида). У эукариот нити ДНК представляют сложные образования, так же называемые хромосомами, которых может быть много на клетку. Хромосомы эукариот отделены от цитоплазмы ядерной мембраной, образуя ядро. Цитологическое различие в строении генетического аппарата является основным в разграничении прокариот и эукариот, остальные признаки служат либо дополнительными, либо коррелирующими. Помимо хромосомы, у прокариот могут присутствовать небольшие кольцевые молекулы ДНК, называемые плазмидами, которые не являются обязательными для генома данного вида. Плазмиды варьируют по величине и по численности в клетке от одной до многих десятков. Крупные плазмиды могут содержать сотни генов. Плазмиды относятся к экстрахромосомным носителям генетической информации.
Нуклеиновые кислоты представляют простые молекулы в виде нити из пентоз (дезоксирибоз для ДНК и рибоз для РНК), соединенных фосфодиэфирной связью. К остаткам пентоз присоединены азотистые основания - пурины, аденин (А) и гуанин (Г), и пиримидины, цитозин (Ц) и тимин (Т). Нить может оканчиваться либо фосфатом (5' конец), либо пентозой (3' конец).
Согласно правилу Чаргаффа, отношение между основаниями в ДНК строго стехиометрическое, но число пар Г + Ц или А + Т может варьировать. По числу преобладания тех или иных пар различают организмы с высоким или низким молярным содержанием ГЦ (ГЦ, моль %), величина которого обычно приводится
58
для вида. Молекула ДНК представляет двойную нить, в которой пары оснований располагаются строго друг против друга (А:Т, Г:Ц) и связаны водородными связями, приводящими к строению молекулы в виде "двойной спирали Уотсона-Крика". Генетическая информация определяется линейным порядком расположения оснований, их последовательностью. Техническая операция установления последовательности называется секвенированием и представляет сейчас важнейший технический метод молекулярной биологии. Геном содержит исчерпывающую характеристику организма и для бактерий имеет величину n·106 пар оснований (п. о.), в типичном случае около 5 млн п. о. Общая длина хромосомы составляет примерно 1 мм и, чтобы упаковать ее в нуклеоид размерами 0,1-0,5 мкм двойная нить сворачивается определенным образом.
Репликация ДНК представляет полуконсервативный процесс, в котором дочерние клетки получают по одной нити материнской и одной нити вновь синтезированной ДНК. Репликация хромосомы начинается в участке, называемом оriС (от англ, origin), присоединением белковых факторов инициации. Это сопровождается расплетением двойной спирали ДНК с образованием двух репликативных вилок, на каждой из которых начинается встречный синтез второй нити. Для синтеза существует сложный ферментный аппарат, обеспечивающий расплетение нитей (ДНК-гираза или топоизомераза), хеликаза, праймаза с праймером РНК, ДНК-полимераза I и ДНК-полимераза III. Их комплекс получил название реплисомы (рис. 20, 21). Для ее функционирования необходимо поступление предшественников в виде праймеров РНК, в свою очередь образуемых из пентоз, оснований, АТФ - из цитоплазмы и ферментов - из рибосомального аппарата.
Следующий этап представляет коррекция считывания, заключающаяся в удалении ошибочно вставленных оснований и замене их на правильные, а также модификация ряда оснований. Материнская нить ДНК имеет метилированные основания, а вновь синтезированная - нет. Коррекцию осуществляют ДНК-полимеразы I и III. Точность репликации ДНК очень высока и ошибки составляют менее 10-9. Для устранения повреждений в результате внешних воздействий, например радиационных, существуют системы репарации.
59
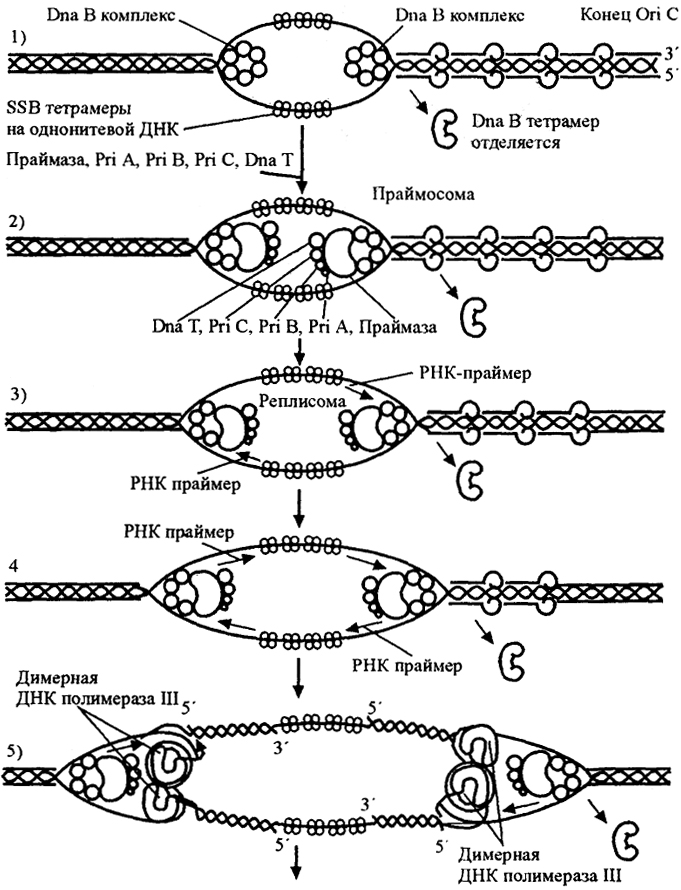
Рис.20. Реплисома (по J. Perry, J. Staley, 1997)
Образование на двойной нити кольцевой хромосомы прокариот двойной вилки включает следующие этапы: 1) - удаление тетрамеров белка Dna В с дуплекса ДНК у конца места начала репликации
Ori С; 2) в месте вилки симметрично присоединяется праймаза с несколькими белками
(Pri A,
Pri В,
Рп С,
Dna Т); 3, 4) праймер РНК синтезируется праймазой для каждой нити ДНК; 5) - присоединяется димерная ДНК-полимераза III.
60
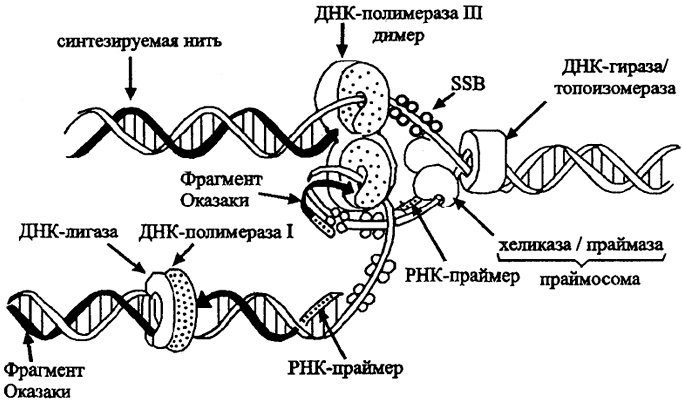
Рис.21. Действие реплисомы (по J. Perry, J. Staley, 1997)
Дуплекс ДНК расплетается ДНК гиразой и хеликазой. Однонитевые участки покрываются дестабилизирующими SSB-белками. В промежуток добавляются праймеры РНК. Димерная ДНК полимераза III связана с нитью скользящим затвором и добавляет нуклеотиды к 3' концу ведущей нити. К ведомой нити добавляется фрагмент Оказаки. ДНК полимераза I и ДНК-лигаза удаляют праймеры, заменяя из соответствующими нуклеотидами.
Наиболее общим механизмом исправления ошибок служит вырезание поврежденного участка и репарация пробела комплементарно сохранившейся нити. Эндонуклеаза гидролизует фосфатные связи на несущей мутацию нити, а затем ДНК-полимераза I восстанавливает пробел. Распознавание поврежденных участков осуществляет SOS-система, вступающая в действие, когда вилка репликации попадает на поврежденное место.
Для ряда бактерий, преимущественно патогенных, составлены полные кольцевые карты ДНК с указанием расположения на них генов. Наиболее подробно изучена Escherichia coli, с которой сравнивают обычно все остальные бактерии. Важно отметить, что последовательности нуклеотидов дают точную характеристику вида организма для его идентификации, сравнимую с номером паспорта, но содержащую также и всю функциональную информацию, которую, однако, нужно уметь прочитать.
61
Одной из важнейших функций клетки служит защита от инородной информации. Для этого клетка обладает мощной и очень активной ДНКазой, гидролизующей инородную ДНК, не находящуюся в клетке в виде кольцевых молекул. Обмен генетической информацией предполагает обходные пути для преодоления этого барьера.
62
