Обмен клетки направлен на поддержание и умножение веществ, из которых состоит клетка. Для этого необходимы энергия и строительные блоки. И то и другое получается в результате переработки поступающих извне веществ внутри клетки. Энергетический обмен, или катаболизм, ведет к получению энергии, а конструктивный обмен, или анаболизм - к образованию строительных блоков. Они составляют серии последовательных реакций, образующих пути метаболизма и объединенных в метаболическую сеть. Катаболизм и анаболизм в той или иной степени сопряжены, и общую их часть иногда называют амфиболизмом или центральными метаболическими реакциями. Следует заметить, что специальные пути катаболизма свойственны литотрофньш микроорганизмам, окисляющим неорганические вещества. В этом случае промежуточные метаболиты не обязательно принадлежат амфиболическим реакциям.
Метаболизм бактерий определяет "типы обмена" или "трофии" - основу группирования их по "физиологическим группам" (см. часть 2). Каждый тип питания характеризуется определенными
41
путями обмена и соответствующими ключевыми ферментами. Пути метаболизма составляют существенную часть биохимии. Здесь нет возможности излагать их, и придется ограничиться лишь кратким перечислением и ролью в системе. На русский язык переведены краткие учебники Г. Шлегеля "Общая микробиология" и Г. Готтшалка "Метаболизм бактерий", сконцентрированные на обмене бактерий, где этот аспект изложен достаточно подробно и ясно для неспециалиста.
Среди процессов катаболизма, осуществляемых микроорганизмами, первостепенное значение имеет разложение углеводов (глюкозы). Наиболее распространенным является путь Эмбдена-Мейергофа, или гликолиз (рис. 9), в котором при окислении одной молекулы глюкозы образуются в конечном итоге две молекулы пирувата, выигрываются две молекулы АТФ и две молекулы восстановителя (НАДН). Такой путь характерен для многих анаэробных органотрофных сахаролитических организмов, осуществляющих брожение углеводов. Проблему для анаэробов представляет удаление избытка восстановителя и регенерация НАД как окислителя. Обычно акцепторами служат органические соединения, в результате их восстановления образуются летучие жирные кислоты (ЛЖК), спирты и Н2. По набору образуемых продуктов различают спиртовое, маслянокислое, молочнокислое, пропионовое, смешанное брожения (рис. 10) и соответствующие функциональные группы анаэробов.
Многие бактерии окисляют глюкозу и по другому пути, Энтнера-Дудорова, с промежуточным образованием 2-кето-З-дезок-си-6-фосфоглкжоновой кислоты. Итогом являются 2 молекулы пирувата, 2 молекулы НАДН и АТФ.
Наконец, разложение глюкозы через пентозофосфатный окислительный цикл включает ряд реакций окисления и декарбокси-лирования глюкозо-6-фосфата с образованием рибулозо-5-фосфата и СО2. Далее следует серия превращений фосфатов Сахаров и регенерация глюкозо-6-фосфата. Одним из интермедиатов пути является 3-фосфоглицерат, который легко может быть превращен в пируват. Пентозофосфатный цикл имеет ряд сходных ступеней с циклом Кальвина - циклическим путем фиксации СО2 у большинства автотрофов, а также с путем ассимиляции формальдегида
42
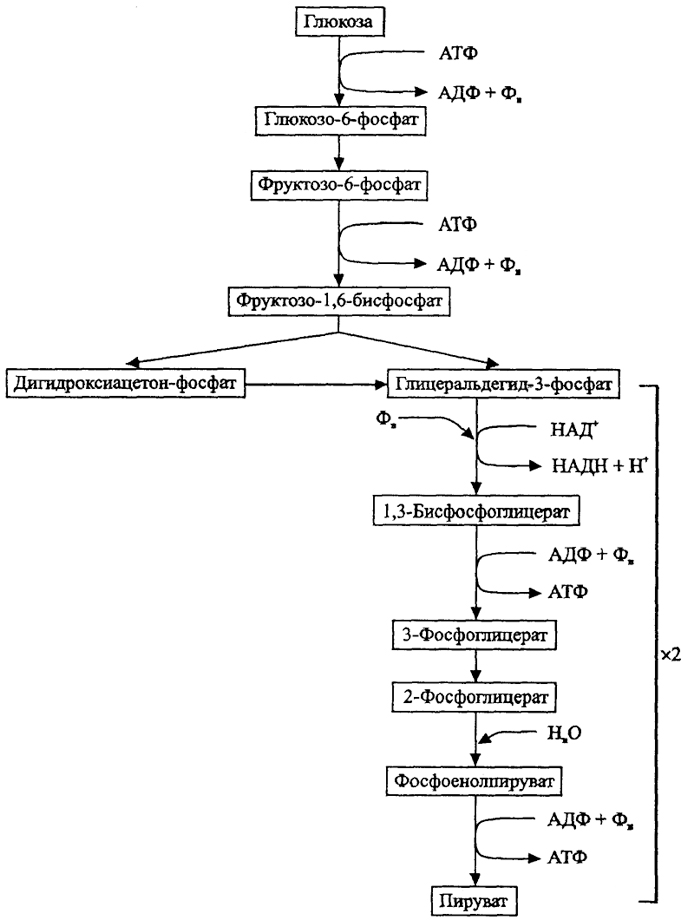
Рис. 9. Путь Эмбдена-Мейергофа (гликолиз) (по Г. Шлегелю,1987)
у метилотрофов. Это обеспечивается полной обратимостью реакций между фосфосахарами, находящихся в равновесии между собой, благодаря действию ферментов трансальдолазы и транске-толазы.
43
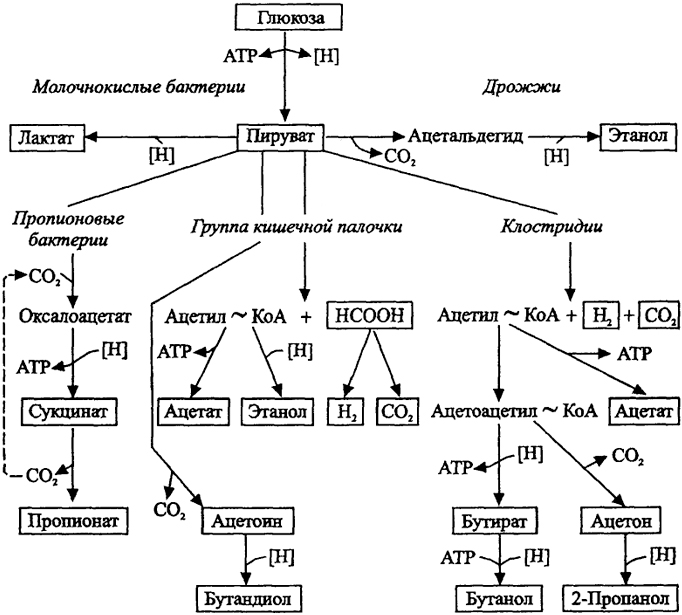
Рис.10. Продукты брожения глюкозы у важнейших групп организмов-бродильщиков (по Г. Шлегелю, 1987)
Пируват, образованный в реакциях катаболизма глюкозы, является исходным компонентом центрального метаболизма. При его окислительном декарбоксилировании образуется ацетил-ко-фермент А (ацетил-КоА). Он может вступать в цикл трикарбоновых кислот (ЦТК), называемый иначе циклом Кребса (рис. 11), в котором последовательные реакции дегидрогенизации и декар-боксилирования составляют циклическую серию превращений три- и дикарбоновых кислот. Действуя в катаболическом направлении, ЦТК ведет к разложению ацетата на 2 молекулы СО2 и 4 молекулы восстановителя, НАД(Ф)Н и ФАДН, которые могут затем окисляться в дыхательной цепи с образованием АТФ. Конечным акцептором электронов может служить O2 или при анаэробном
44

Рис.11. Цикл трикарбоновых кислот (ЦТК)
дыхании, свойственном вторичным анаэробам, другие неорганические и органические вещества. Окисление при этом может идти до конца или же быть неполным. Значительное число веществ, используемых микроорганизмами, может быть превращено в ацетат и, следовательно, вступать в ЦТК и другие важнейшие метаболические пути.
Промежуточные продукты центрального метаболизма служат исходными для синтеза строительных блоков в реакциях
45
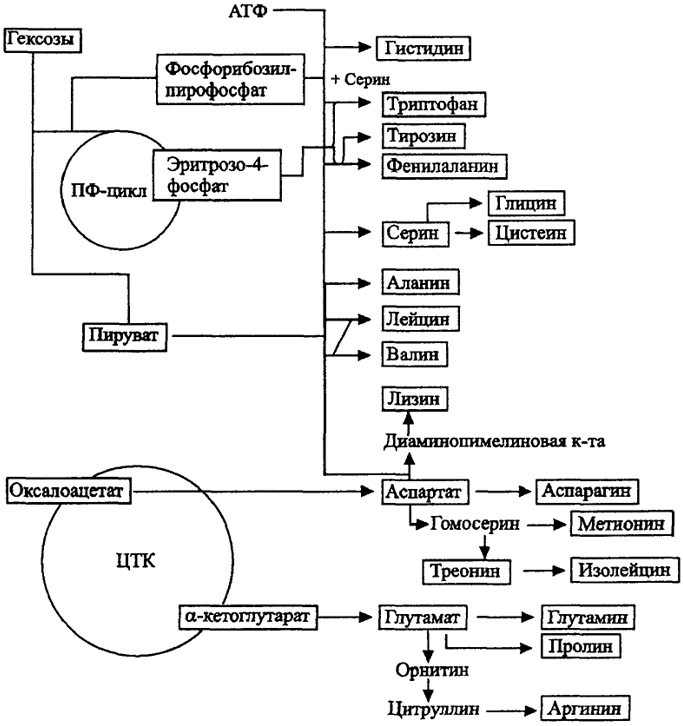
Рис. 12. Основные пути биосинтеза аминокислот (по Г. Шлегелю, 1987) ПФ - пентозофосфатный окислительный путь; ЦТК - цикл трикарбоновых кислот
анаболизма. Так, большинство углеродных скелетов аминокислот для синтеза белков поставляет ЦТК. Для синтеза 20 аминокислот достаточно всего 6 исходных метаболитов (рис. 12). Другие пути ведут к пуринам, пиримидинам, липидам, фосфолипидам, углеводам и всем другим компонентам клетки, образуя сложную транспортную сеть веществ (рис. 13). В этой метаболической сети важная регуляторная функция принадлежит концентрации метаболитов.
46

Рис.13. Взаимосвязь путей катаболизма и анаболизма (по G.Tortora et al., 1998) Двойными стрелками показаны амфиболические пути
Из строительных блоков, представляющих активированные формы метаболитов в виде фосфатных эфиров, аденйлатов, соединений, связанных с коферментом А, образуются макромолекулы. Среди них выделяют необходимые, как нуклеиновые кислоты, белки, фосфолипиды мембраны, компоненты клеточной стенки, и необязательные, как полимерные запасные соединения
47
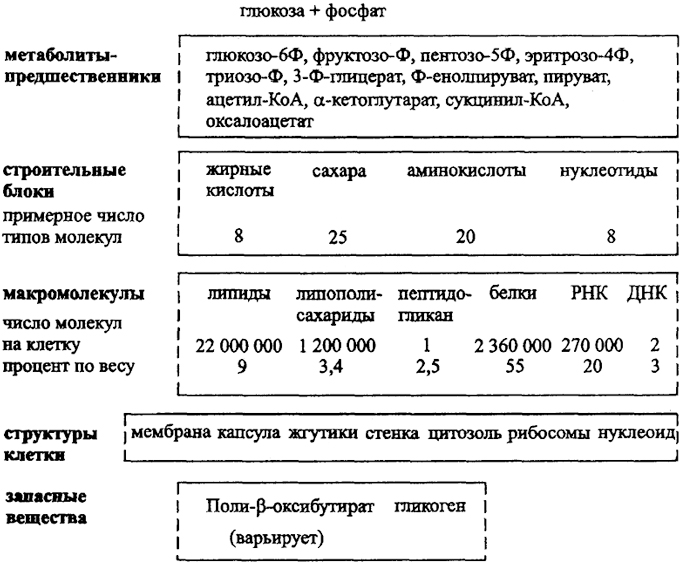
Рис. 14. Общая схема анаболизма хемоорганогетеротрофа
или вещества капсулы (рис. 14). Запасные полисахариды, характерные для организмов, способных к их анаэробному использованию, образуются путем глюконеогенеза. Синтез липидов, в том числе характерного для многих бактерий поли-р-оксибутирата, осуществляется с использованием ацетил-КоА.
Развитые реакции центрального метаболизма позволяют организму осуществлять биосинтетические функции за счет единственного, иногда даже необычного по строению органического вещества. Напротив, яркий случай гетеротрофии представляют организмы, нуждающиеся для биосинтеза во многих предшественниках. Примером может служить молочнокислая бактерия Leuconostoc mesenteroides, для роста которой в синтетическую среду следует добавлять 19 аминокислот и 4 пурина и пиримидина. При культивировании гетеротрофных микроорганизмов сумма возможно необходимых строительных блоков заменяется внесением
48
в среду дрожжевого экстракта и набора витаминов. Особенно сложными Пищевыми потребностями обладают паразитические микроорганизмы, часть из которых вообще не растет вне хозяина.
Чтобы оказаться совместимыми с реакциями центрального метаболизма, вещества, потребляемые клеткой в качестве субстратов катаболизма, должны претерпеть ряд предварительных превращений, своих для каждого соединения. Эти реакции, иногда называемые подготовительным метаболизмом, у бактерий чрезвычайно разнообразны.
Возможность использования субстратов микроорганизмами определяется способом превращения их в транспортируемые в клетку вещества и от совместимости с имеющимся в клетке ферментным аппаратом. Полисахариды гидролизуются экзоферментами до дисахаридов, при этом легко гидролизуются растворимые полисахариды, как гликоген или крахмал, и труднее нерастворимые, как целлюлоза. Дисахариды обычно разлагаются на моносахариды. Гексозы используются многими организмами и для этого имеются специальные транспортные системы. Пентозы обычно трансформируются в гексозы и глицеральдегид-фосфат. Набор углеводов, используемых микроорганизмами, видоспецифичен и особенно широко применялся для идентификации энтеробактерий.
Белки гидролизуются в пептиды и далее в аминокислоты, поступающие в клетку, где они дезаминируются и превращаются в органические кислоты. Однако часть аминокислот, например аргинин, разлагается своими путями. У анаэробных протеолитических организмов имеются различные пути брожения аминокислот и других органический соединений азота. Конечными продуктами здесь являются аммиак, жирные кислоты, обычно разветвленные, и ацетат.
На органических кислотах многие аэробные бактерии растут очень быстро и охотно. Основным путем их превращений является ЦТК, причем возможность использования в реакциях и катаболизма, и анаболизма обеспечивается дополнительными (анаплеротическими) метаболическими путями, ключевая роль в которых принадлежит С2- соединениям, как ацетил-КоА. Рост на среде с ацетатом может осуществляться за счет ЦТК и реакций глиок-силатного цикла с ключевыми ферментами малатсинтазой и
49
изоцитратлиазой. Этот путь является основным для использования липидов и углеводородов, где ацетил-КоА образуется в реакциях Р-окисления. Возможность расти на глйоксилате, продукте разложения пуринов, обеспечивается его превращением в ацетил-КоА в гликолатном пути.
Одноуглеродные соединения не могут быть использованы обычным путем, и для перехода к анаболизму требуется превращение их в многоуглеродные соединения, т.е. синтез С-С связи. Поэтому к использованию С1- соединений способны только специалисты. Восстановленные С1 -соединения, как метан и метанол, включаются в обмен после окисления в формальдегид. Последний может вступить в центральный метаболизм либо через рибулозомонофосфатный цикл с образованием гексозо-6-фосфа-та, либо через сериновый путь с фиксацией формальдегида на глицине и образованием серина. Оксид углерода служит субстратом для карбоксидобактерий, которые окисляют его в диоксид. СО может также включаться в обмен через СО-дегидрогеназу и вступать в путь Льюнгдала-Вуда, или ацетил-КоА путь, свойственный анаэробным гомоацетатным бактериям. У ряда прокариот восстановительный ацетил-КоА путь обеспечивает авто-трофную ассимиляцию углекислоты в результате образования из двух молекул СО2 молекулы ацетата.
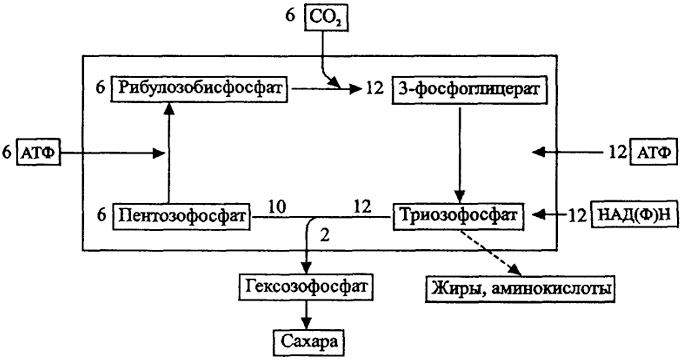
Рис. 15. Схема автотрофной ассимиляции СО (цикл Кальвина)
50
Подавляющая часть автотрофных организмов ассимилирует СO2 по восстановительному пентозофосфатному пути (циклу Кальвина) с ключевым ферментом рибулозобисфосфат карбокси-лазой (рубиско). Этот путь (рис. 15) включает карбоксилирование рибулозобисфосфата с образованием двух молекул трехуглеродного фосфоглицерата и последующие циклические превращения фосфатов Сахаров с регенерацией рибулозобисфосфата, и в конечном итоге выходом из цикла молекулы глюкозы. Цикл Кальвина работает с потреблением 18АТФ и 12НАДФН :
6СО2 + 12НАДФН + 18АТФ - С6Н12О6 + 12НАДФ + 18АДФ +18 Фнсорг
Некоторые автотрофы, например Hydrogenobacter (Caldero-bacterium) или зеленая серная бактерия Chlorobium, используют для автотрофной ассимиляции СО2 восстановительный ЦТК. У автотрофных организмов анаболические реакции отделены от катаболических. Чтобы блокировать сгорание органического вещества, ЦТК у автотрофов часто бывает разорван и служит лишь для синтеза предшественников.
Суммируя, надо сказать, что каждая функциональная группа бактерий может быть охарактеризована свойственным ей путем обмена. В свою очередь, для каждого пути обмена существуют характерные ферменты или комбинации ферментов, измеряя активность которых можно определить функционирование этого пути. Ферменты представляют собой специфические белки, структура которых кодирована в ДНК определенной последовательностью нуклеотидов, составляющих ген. Отсюда есть возможность определить потенциальную способность к осуществлению тех или иных реакций по анализу ДНК в чистой культуре или даже в суммарной ДНК сообщества. Такой подход сейчас широко развивается на основе генных проб на определенные гены.
С другой стороны, значительная часть метаболических путей, в особенности направленных на биосинтетические нужды, универсальна и соответствующие ферменты и гены могут быть обнаружены у широкого круга организмов.
Блок цитоплазмы связан с генетическим аппаратом клетки по средством регуляторных и/или сигнальных путей, которые определяют экспрессию синтеза ферментов в процессе адаптации. Блок цитоплазмы связан с мембранным аппаратом, а через него с внешней средой - процессами транспорта.
51
